

Energy metabolism factors in umbilical cord blood in fetal growth restriction
Kan N.E., Tyutyunnik V.L., Soldatova E.E., Ryzhova K.O., Gaidarova A.R., Krasnyi A.M.
Objective: To investigate the concentrations of energy metabolism factors in umbilical cord blood plasma in fetal growth restriction (FGR).
Materials and methods: A case-control study was conducted involving 118 patients. The study group comprised 60 pregnant women with a postnatally confirmed diagnosis of FGR, while the comparison group comprised 58 women with physiologically uncomplicated pregnancies without FGR. The concentrations of energy metabolism factors (C-peptide, ghrelin, glucose-dependent insulinotropic polypeptide, glucagon-like peptide-1, glucagon, insulin, leptin, plasminogen activator inhibitor-1, resistin, and visfatin) in umbilical cord blood plasma were determined using multiplex analysis with a 10-plex Bio-Plex Pro Human Diabetes Panel assay system.
Results: Statistically significant differences were identified in the concentrations of ghrelin, leptin, and plasminogen activator inhibitor-1 (PAI-1) (p<0.001) in umbilical cord blood plasma. In the FGR group, ghrelin (221.89 [160.08; 281.71] ng/mL) and PAI-1 (2150.48 [1604.57; 2859.83] ng/mL) concentrations were significantly elevated, whereas leptin concentrations were reduced (213.58 [101.57; 1027.24] ng/mL). Furthermore, a moderate positive correlation was established between low anthropometric indices of the neonate and leptin concentrations (rs=0.34, p=0.04 and 0.46, p<0.01), and a negative correlation with ghrelin concentrations
(rs=-0.51, p=0.003 and -0.52, p<0.01) in patients with FGR.
Conclusion: These findings indicate that energy metabolism factors are implicated in the pathogenesis of FGR and warrant further investigation. FGR is characterized by reduced leptin and elevated ghrelin concentrations, reflecting the involvement of the growth hormone–insulin-like growth factor-1 axis in the pathogenesis of this obstetric complication. Dysregulation of these hormones and adipokines in neonates may be a risk factor for the development of metabolic disorders in adulthood.
Authors' contributions: Kan N.E., Tyutyunnik V.L., Soldatova E.E., Ryzhova K.O., Gaidarova A.R., Krasnyi A.M. – conception and design of the study, data collection, literature review, processing and analysis of relevant material, drafting of the manuscript, editing of the manuscript.
Conflicts of interest: The authors have no conflicts of interest to declare.
Funding: There was no funding for this study.
Ethical Approval: The study was reviewed and approved by the Research Ethics Committee of the V.I. Kulakov NMRC for OG&P, Ministry of Health of Russia.
Use of generative artificial intelligence: none.
Patient Consent for Publication: All patients provided informed consent for the publication of their data.
Authors' Data Sharing Statement: The data supporting the findings of this study are available upon request from the corresponding author after approval from the principal investigator.
For citation: Kan N.E., Tyutyunnik V.L., Soldatova E.E., Ryzhova K.O., Gaidarova A.R., Krasnyi A.M.
Energy metabolism factors in umbilical cord blood in fetal growth restriction.
Akusherstvo i Ginekologiya/Obstetrics and Gynecology. 2026; (4): 77-86 (in Russian)
https://dx.doi.org/10.18565/aig.2025.365
Keywords
fetal growth restriction
umbilical cord blood
ghrelin
leptin
plasminogen activator inhibitor-1 (PAI-1)
fetal programming
Fetal growth restriction (FGR) remains highly relevant in obstetrics, representing one of the principal causes of adverse perinatal outcomes and contributing substantially to perinatal and infant morbidity and mortality [1, 2].
Adverse exposures affecting the maternal organism during pregnancy initiate a cascade of epigenetic alterations in fetuses. This process, known as fetal programming, lays the groundwork for the future development of metabolic syndrome, cardiovascular diseases, and neurological disorders [3–5]. The principal explanatory framework is David Barker's "thrifty phenotype" hypothesis, according to which the fetus, under conditions of nutrient deficiency and chronic hypoxia, redistributes energy resources through reduced tissue sensitivity to fetal insulin (insulin resistance) and alterations in glucose homeostasis [6].
Adipokines, hormonally active substances produced by adipose tissue, play a critical role in the mechanisms of intrauterine programming. Among their considerable diversity, leptin and adiponectin are of particular significance in predicting future metabolic disturbances [7, 8]. The concentrations of these biomarkers in neonates correlate directly with the volume of adipose tissue formed by the fetus at the time of delivery. Consequently, the analysis of the adipokine profile at birth provides objective information regarding the degree of adipose tissue development and may serve as an early indicator of potential individual metabolic risks [8].
In addition to adipokines, other hormonal factors play important roles in regulating fetal growth processes. The principal hormones involved in this process are insulin, secreted by the β-cells of the pancreatic islets of Langerhans, and ghrelin, produced predominantly by the enteroendocrine cells of the gastrointestinal mucosa [9]. The functional interaction of these biologically active substances is mediated, in part, through modulation of the somatotropic growth hormone–insulin-like growth factor-1 (GH–IGF-1) axis, which directly influences the rate of intrauterine development. An imbalance within this hormonal system, induced by various prenatal factors, may be a potential predictor of metabolic disturbances in the postnatal period [10].
From this perspective, the investigation of the concentrations of factors that exert a substantial influence on the regulation of energy balance and glucose metabolism is of interest, with a view to clarifying their role in intrauterine fetal development.
This study aimed to investigate the concentrations of energy metabolism factors in umbilical cord blood plasma in cases of fetal growth restriction.
Materials and methods
A case-control study was conducted that enrolled 118 pregnant women. The study group (Group I) comprised 60 patients with a postnatally confirmed diagnosis of FGR, and the control group (Group II) comprised 58 patients with uncomplicated singleton pregnancies.
The diagnosis of FGR during pregnancy was established in accordance with the clinical guidelines "Insufficient fetal growth requiring maternal medical care (fetal growth restriction)" [11]. Neonatal anthropometric parameters (birth weight, body length, and head circumference) were assessed using INTERGROWTH-21 percentile tables adapted for different gestational ages [12]. Postnatal confirmation of the diagnosis of FGR was based on the criteria of the International Consensus (2018). The diagnosis was considered confirmed when neonatal weight-for-length indices fell below the 3rd percentile or when at least three of the following criteria were present: birth weight below the 10th percentile, head circumference below the 10th percentile, body length below the 10th percentile, prenatally diagnosed FGR, and complicated maternal pregnancy (arterial hypertension or pre-eclampsia) [13].
The study was reviewed and approved by the Research Ethics Committee of V.I. Kulakov National Medical Research Centre for Obstetrics, Gynecology, and Perinatology of the Ministry of Health of the Russian Federation. All participants provided written informed consent prior to enrolment.
The inclusion criteria were as follows: for the study group, singleton pregnancy complicated by FGR; for the control group, physiologically uncomplicated singleton pregnancy without signs of FGR. All participants were required to sign an informed consent form.
Patients were excluded from the study if any of the following criteria were present (non-inclusion criteria): multiple pregnancies, development of pre-eclampsia during pregnancy, use of donor oocytes in in vitro fertilization (IVF) programs, severe extragenital pathology, antiphospholipid syndrome, acute infectious disease during gestation, confirmed fetal malformations, or diagnosed hemolytic disease of the fetus and newborn.
The quantitative determination of the energy metabolism biomarker panel (C-peptide, ghrelin, glucose-dependent insulinotropic polypeptide [GIP], glucagon-like peptide-1 [GLP-1], glucagon, insulin, leptin, plasminogen activator inhibitor-1 [PAI-1], resistin, and visfatin) in umbilical cord blood was performed using multiplex analysis. Venous blood samples (5 mL) were collected from umbilical cord vessels immediately after delivery into vacuum collection tubes containing ethylenediaminetetraacetic acid (EDTA). Pre-analytical sample preparation was completed within 30 min of collection. Plasma separation was performed by double centrifugation (500 × g and 5000 × g, 10 min each). The resulting 500 µL aliquots were cryopreserved at -80°C [14].
Analyte concentrations were measured using the commercial Bio-Plex Pro Human Diabetes Panel (10-plex, Bio-Rad, Hercules, California, USA) in strict accordance with the manufacturer's instructions on the Bio-Plex 200 hardware-software platform (Bio-Rad, USA). Primary data processing was performed using the Bio-Plex Manager 6.0 software (Bio-Rad, USA). Analyte concentrations were reported in ng/mL [14].
Statistical analysis
Statistical analysis was performed using the following software packages: Microsoft Excel, OriginPro 8, SPSS Statistics 17, and StatTech v. 3.1.4 (developed by StatTech LLC, Russia). As the data distribution deviated from normality, the Mann–Whitney U test was used to compare independent samples. Descriptive statistics are presented as medians (Me) with lower and upper quartiles (Q1; Q3). Data are displayed graphically as box-and-whisker plots reflecting percentile distributions (5th, 25th, 50th, 75th, and 95th). For the analysis of categorical variables, Fisher's exact test was applied, the and results are presented as counts and proportions (n/N, %). The associations between variables were assessed using Spearman's rank correlation coefficient (rs). Differences and associations were considered statistically significant at p<0.05.
Results and discussion
As is well known, energy metabolism during pregnancy is regulated through bidirectional exchange within the maternal-fetal system. In this regard, we analyzed the baseline clinical and anamnestic characteristics of the women, as well as the course of pregnancy and delivery (Table 1).
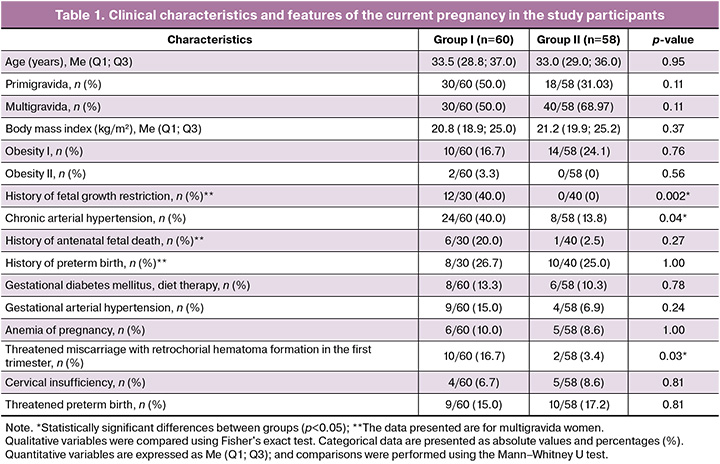
Analysis of anthropometric parameters (body weight, height, and body mass index (BMI)) and age revealed no significant differences between the groups (p>0.05). The median age of pregnant women in the study group was 33.5 years [28.8; 37.0], while in the control group it was 33.0 years [29.0; 36.0] (p=0.95). Evaluation of somatic status demonstrated significant differences in the prevalence of chronic arterial hypertension (p=0.04), which was more common in patients of the study group (24/60, 40%) than in the control group (8/58, 13.8%).
Analysis of obstetric history showed that among multigravida women, fetal growth restriction (FGR) in previous pregnancies was significantly more frequent in the study group (12/30, 40%; p=0.002). In addition, the current pregnancy in this group was more often complicated by threatened miscarriage with the formation of a retrochorial hematoma in the first trimester (10/60, 16.7%; p=0.03).
The median gestational age at delivery in the study group was 37.3 (35.4; 38.1) weeks, compared with 38.3 (37.0; 39.0) weeks in the control group (p=0.002). In 31.7% (19/60) of the cases, pregnancy ended in spontaneous vaginal delivery (p=0.09), whereas 68.3% (41/60) of the patients underwent cesarean section (p=0.09). Among the pregnant women with FGR, 33.3% (20/60) had preterm deliveries. The main indications for cesarean section were deterioration of fetal condition according to functional diagnostic methods and lack of fetal growth dynamics on ultrasonography.
A comparison of placental morphometric characteristics revealed statistically significant differences between the groups (p<0.05). In the study group, placental weight and umbilical cord length were significantly lower (p<0.001), amounting to 268 (200; 312) g and 24 (18; 33.3) cm, respectively, compared with 450 (384; 511) g and 35.5 (24.5; 43.5) cm in the control group. Marginal cord insertion was observed in 53.3% of cases, velamentous insertion in 3.3%, eccentric insertion in 36.7%, and central insertion in only 6.7% (p<0.001).
The Apgar scores of newborns ranged from 5 to 9 (p=0.02). According to the INTERGROWTH-21 percentile charts, the neonatal anthropometric parameters were significantly lower in the study group than in the control group (Table 2).
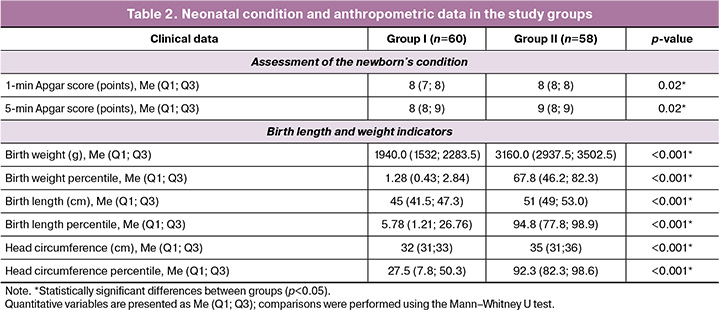
The birth weight in the study group was 1940.0 (1532; 2283.5) g versus 3160.0 (2937.5; 3502.5) g in the control group (p<0.001). The birth weight percentiles were 1.28 (0.43; 2.84) and 67.8 (46.2; 82.3), respectively (p<0.001). Neonatal length was 45.0 (41.5; 47.3) cm in the study group and 51.0 (49.0; 53.0) cm in the control group (p<0.001). Length percentiles were also significantly lower in FGR newborns (5.78 (1.21; 26.76) vs. 94.8 (77.8; 98.9), respectively (p<0.001).
The early neonatal period was more frequently complicated in newborns in the study group (Fig. 1).
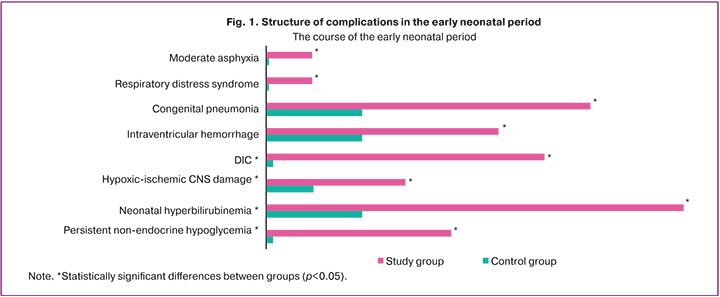
Among respiratory disorders, moderate birth asphyxia was observed in 10% (6/60) of cases (p=0.03), respiratory distress syndrome in 11.7% (7/60) (p=0.01), and congenital pneumonia in 23.3% (14/60) (p=0.02) of cases.
Hypoxic–ischemic central nervous system (CNS) injury was significantly more frequent in the study group (6/60, 10.0%; p=0.03). Intraventricular hemorrhage was detected by neurosonography in 16.7% (10/60) of the FGR newborns (p=0.03). Disseminated intravascular coagulation syndrome occurred in 20% (12/60) of the patients (p=0.001).
Persistent hypoglycemia (8/60, 13.3%; p=0.006) and congenital hyperinsulinism (4/60, 6.7%; p=0.12) were also more common in FGR newborns. However, glucose levels did not differ significantly between the groups (p=0.87), being 3 mmol/L (2; 3.43) in the study group and 3.2 mmol/L (2.58; 3.45) in the control group.
Birth weight and length reflect intrauterine development and are essential tools for the clinical assessment of neonatal conditions. These parameters are routinely used in clinical practice to identify newborns at risk of complications, both in the neonatal period and later in life.
Impaired fetal growth is associated with multiple factors, including hormonal influences, particularly those that regulate energy metabolism. However, the mechanisms linking fetal growth and metabolic disturbances remain unclear. In our previous studies [14, 15], we described the levels of hormones, incretins, and adipokines in maternal plasma, as well as the gene expression involved in energy metabolism regulation in maternal blood, cord blood, and placental tissue in FGR. These findings helped clarify the potential role of energy metabolism factors and their regulatory genes in the pathogenesis of this pregnancy complication from the perspective of fetal programming.
Given the complex interactions within the mother–placenta–fetus system, we investigated the energy metabolism factors in umbilical cord plasma (Table 3). The results have been published and defended as part of a dissertation by E.E. Soldatova entitled: "Clinical and pathogenetic features of fetal growth restriction based on the assessment of energy metabolism factors and the expression of their genes."
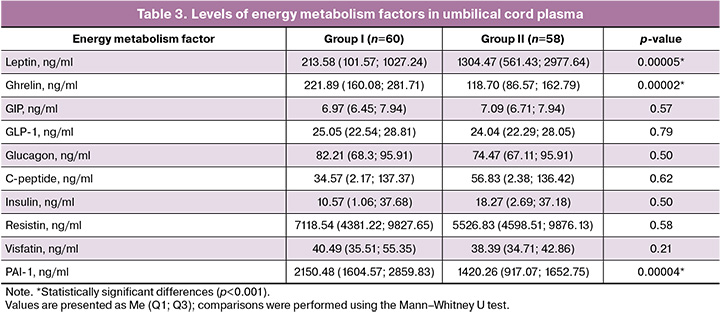
Comparative analysis revealed statistically significant differences in three parameters: ghrelin, leptin, and plasminogen activator inhibitor-1 (PAI-1) (p<0.001).
Ghrelin levels were significantly increased (p<0.001) in the cord plasma of FGR newborns compared to normal-weight neonates: 221.89 (160.08; 281.71) ng/mL versus 118.70 (86.57; 162.79) ng/mL.
In contrast, the counter-regulatory adipokine leptin was significantly decreased in FGR newborns (p<0.001), measuring 213.58 (101.57; 1027.24) ng/mL versus 1304.47 (561.43; 2977.64) ng/mL in the control group, respectively.
PAI-1 levels also differed significantly (p<0.001), being nearly twice as high in the study group (2150.48 [1604.57; 2859.83] ng/mL) than in the control group 1420.26 (917.07; 1652.75] ng/mL). Elevated PAI-1 levels in cord blood may reflect chronic intrauterine hypoxia in FGR, which activates HIF-1α and increases SERPINE1 gene expression in the vascular endothelium, placenta, and fetal tissues. This leads to enhanced PAI-1 synthesis and secretion into fetal circulation, contributing to endothelial dysfunction, a procoagulant state, inflammation, and the development of insulin and leptin resistance in the fetus [16]. Thus, elevated PAI-1 links two key pathological processes: impaired placental hemostasis and fetal insulin resistance. It may serve as an early marker of a pathological metabolic profile associated with metabolic syndrome, potentially predisposing individuals to disease in adulthood [17].
No statistically significant differences were observed for other factors (Table 3, Fig. 2).
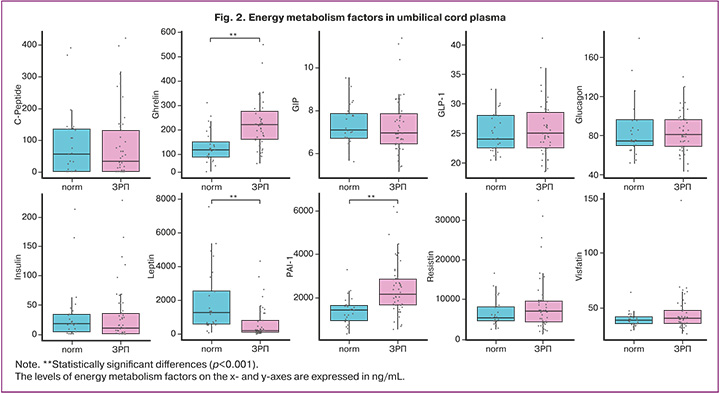
According to the literature, leptin and ghrelin levels in cord blood correlate with neonatal anthropometric parameters at birth [18, 19]. In this study, a correlation analysis was performed to assess the relationship between these hormones and neonatal weight and length (Fig. 3).
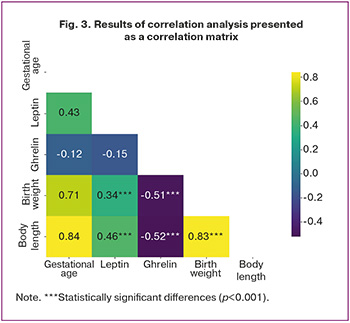
In FGR newborns, leptin levels showed a moderate positive correlation with birth weight (Spearman's rs=0.34; p=0.04) and birth length (rs=0.46; p<0.01). No significant correlations were observed in the control group (rs=0.36, p=0.38 and rs=0.33, p=0.35, respectively). These findings suggest that leptin may not only be associated with anthropometric parameters but also influence fetal and neonatal metabolism, reflecting not merely adaptation to intrauterine deprivation but also decompensation and the formation of a pathological metabolic profile. No significant correlation was found between leptin levels and gestational age at delivery (rs=0.43; p=0.17).
Ghrelin levels in FGR newborns demonstrated a moderate negative correlation with birth weight (rs=-0.51; p=0.003) and length (rs=-0.52; p<0.01), indicating that higher ghrelin levels are associated with lower birth weight and length. This suggests a role for ghrelin in FGR pathogenesis. No significant correlations were found in the control group (rs=-0.25, p=0.28 and rs=-0.17, p=0.19, respectively), nor between ghrelin levels and gestational age at delivery (rs=-0.12, p=0.28).
Thus, low ghrelin levels at birth may serve as a marker of the severity of neonatal adaptation to intrauterine conditions and reflect the extent to which the fetus suppresses its growth program for survival.
The molecular and biological mechanisms underlying metabolic programming are not fully understood and require further investigation. Among the key mediators of this process, dysfunction of the hypothalamic-pituitary-adrenal axis and impaired insulin signaling have garnered significant attention. The findings presented here support the hypothesis that suboptimal intrauterine conditions can alter the maturation of hormonal regulatory systems, predisposing individuals to metabolic disorders later in life. Ghrelin and leptin play principal roles in energy balance regulation.
Ghrelin, produced mainly in the stomach but also in the placenta, likely has a key role in intrauterine energy homeostasis. During gestation, the fetal pancreas is the primary source of ghrelin [20]; therefore, its concentration in umbilical cord blood may indicate the functional state of the developing pancreas. Published data indicate that ghrelin concentrations in umbilical cord blood of neonates with low birth weight are significantly higher than those of neonates with normal anthropometric parameters [9, 21]. Furthermore, it has been proposed that neonates with low birth weight and low cord blood ghrelin concentrations exhibit attenuated postnatal weight gain [22]. Several studies have reported elevated ghrelin concentrations in umbilical cord blood of growth-restricted neonates, which correlated inversely with neonatal length and birth weight [18, 19].
Evidence suggests that elevated ghrelin concentrations in small-for-gestational-age neonates may be a compensatory response, leading to increased secretion of growth hormone, which regulates fetal and neonatal growth [20, 21]. Ghrelin stimulates growth hormone secretion (somatotropic [growth-promoting] action) and insulin-like growth factor-1 (IGF-1) [23]. Acting directly via the hypothalamus and pituitary gland, ghrelin initiates a cascade that leads to the production of IGF-1 – the principal driver of cell division and tissue growth. Given that fetal growth restriction is accompanied by chronic energy deficit, and that fetal growth is the most energetically demanding biological process, ghrelin suppresses the growth hormone-IGF-1 axis, reducing somatotropic axis activity and attenuating protein synthesis and cell division, ultimately leading to fetal growth retardation and diminished weight accrual [23, 24].
Elevated ghrelin levels stimulate appetite, resulting in increased food intake in neonates after delivery. Ghrelin concentrations are highest in the early postnatal period, before growth hormone assumes its regulatory role over nutrient intake and neonatal growth; thus, ghrelin's function at this stage is similar to that of growth hormone [25], which may explain ghrelin's potential contribution to postnatal catch-up growth in neonates with prenatal growth restriction [22].
Leptin acts as a counter-regulatory hormone and is an important component of glucose energy homeostasis under physiological conditions. Leptin acts mainly at the level of the hypothalamus, suppressing appetite and initiating catabolic pathways. Peripherally, it inhibits adipose tissue accumulation, thereby maintaining glucose sensitivity, and regulates adipose tissue; accordingly, optimal leptin concentrations throughout development promote optimal growth and maturation of the main metabolic regulatory pathways. Even transient periods of hypo- or hyperleptinemia can cause metabolic dysregulation and the activation of compensatory mechanisms [26].
Our findings, consistent with published data [18, 19], demonstrate that neonates with low weight-for-length indices exhibit reduced leptin concentrations in umbilical cord blood, which appears to be due to diminished fat mass and placental dysfunction in the setting of fetal growth restriction. In addition, a positive correlation was observed between low leptin values and birth weight in growth-restricted neonates.
Current literature addresses the relationship between umbilical cord blood leptin concentrations and the severity of fetal hypoxemia. The leptin gene is sensitive to changes in oxygen tension [27, 28]. Chronic intrauterine hypoxia, which typically accompanies fetal growth restriction, stimulates enhanced leptin production by trophoblast cells. However, despite increased synthesis, growth-restricted neonates show a reduction in the biologically active fraction of leptin in umbilical cord blood, likely because of the high-affinity binding of this adipokine to its receptors. This phenomenon may disrupt the fine regulation of feeding behavior and energy metabolism in the postnatal period, including into adulthood [29].
The results of this study align with this pattern. Among the examined growth-restricted neonates, a paradoxical reduction in cord blood adipokine concentrations was observed against a background of upregulated leptin gene expression in placental tissue [15]. The discordance between increased placental leptin gene expression and reduced cord blood leptin levels may indicate a failure of compensatory mechanisms in fetal growth restriction and the involvement of the JAK/STAT signaling pathway – which plays a pivotal role in mediating the effects of leptin and in the fetal programming of metabolic disorders – in the pathological process [28, 30]. Thus, elevated cord blood leptin in the context of fetal growth restriction does not represent an adaptive response to the developing intrauterine environment, but rather signifies decompensation of the metabolic pathway.
Furthermore, the interrelationship among leptin, insulin, and ghrelin in neonates with birth weight below 2,000 g suggests that the fetal adipo-insular axis at birth may influence the "programming" of satiety and metabolism, thereby determining postnatal weight gain and susceptibility to metabolic disorders after delivery [18]. This may be related to alterations in the adipokine profile of umbilical cord blood and ghrelin levels, which in turn may impair the neonatal metabolic adaptation to extrauterine life. These observations suggest that measuring these hormonal factors in umbilical cord blood reflects the neonate's intrauterine developmental trajectory. Nevertheless, the mechanisms underlying the observed associations require further investigation.
Conclusion
This study identified statistically significant differences in energy metabolism factors between patients with pregnancies complicated by fetal growth restriction and those with physiologically uncomplicated pregnancies. The findings indicate that energy metabolism factors may participate in the pathogenesis of fetal growth restriction and highlight the value of further investigation from the perspective of fetal programming of metabolic disorders later in life.
Specifically, fetal growth restriction is characterized by low leptin and glucose concentrations and elevated ghrelin levels, reflecting the involvement of the growth hormone–IGF-1 axis in the pathogenesis of this obstetric complication. In patients whose neonates demonstrated the lowest weight-for-length parameters, a direct correlation with leptin concentrations and an inverse correlation with ghrelin concentrations were observed.
These data support the hypothesis that adult-onset disease arises from alterations in the maturation of hormonal axes under suboptimal intrauterine conditions. Dysregulation of the aforementioned factors in growth-restricted neonates may establish a foundation for the development of metabolic disease in adulthood, representing a manifestation of the "thrifty phenotype" of fetal programming as conceptualized within the Barker hypothesis.
References
- Якубова Д.И., Игнатко И.В., Меграбян А.Д., Байбулатова Ш.Ш., Кузнецов А.С. Возможности прогнозирования и особенности течения беременности при различных фенотипических формах задержки роста плода. Вопросы гинекологии, акушерства и перинатологии. 2022; 21(6): 35-42. [Yakubova D.I., Ignatko I.V., Megrabyan A.D., Baibulatova Sh.Sh., Kuznetsov A.S. Predictability and characteristics of pregnancy course in different phenotypes of fetal growth restriction. Gynecology, Obstetrics and Perinatology. 2022; 21(6): 35-42 (in Russian)]. https://dx.doi.org/10.20953/1726-1678-2022-6-35-42
- Dessì A., Pravettoni C., Cesare Marincola F., Schirru A., Fanos V. The biomarkers of fetal growth in intrauterine growth retardation and large for gestational age cases: from adipocytokines to a metabolomic all-in-one tool. Expert Rev. Proteomics. 2015; 12(3): 309-16. https://dx.doi.org/10.1586/14789450.2015.1034694
- Marciniak A., Patro-Małysza J., Kimber-Trojnar Ż., Marciniak B., Oleszczuk J., Leszczyńska-Gorzelak B. Fetal programming of the metabolic syndrome. Taiwan. J. Obstet. Gynecol. 2017; 56(2): 133-8. https://dx.doi.org/10.1016/j.tjog.2017.01.001
- Петров Ю.А., Купина А.Д. Фетальное программирование – способ предупреждения заболеваний во взрослом возрасте. Медицинский совет. 2020; 13: 50-56. [Petrov Yu.A., Kupina A.D. Fetal programming – a method for preventing diseases in adulthood. Medical Сouncil. 2020; 13: 50-56 (in Russian)]. https://dx.doi.org/10.21518/2079-701X-2020-13-50-56
- Джобава Э.М. Фетальное программирование. Акушерство и гинекология. 2018; 3: 10-5. [Dzhobava E.M. Fetal programming. Obstetrics and Gynecology. 2018; (3): 10-5 (in Russian)]. https://dx.doi.org/10.18565/aig.2018.3.10-15
- Baker J.L., Olsen L.W., Sorensen T.I. Weight at birth hand all-cause mortality in adulthood. Epidemiology. 2008; 19(2): 197-203. https://dx.doi.org/10.1097/EDE.0b013e31816339c6
- Dessì A., Pravettoni C., Cesare Marincola F., Schirru A., Fanos V. The biomarkers of fetal growth in intrauterine growth retardation and large for gestational age cases: from adipocytokines to a metabolomic all-in-one tool. Expert Rev. Proteomics. 2015; 12(3): 309-16. https://dx.doi.org/10.1586/14789450.2015.1034694
- Сукало А.В., Прилуцкая В.А., Солнцева А.В., Уварова Е.В. Современные представления о роли адипоцитокинов в программировании гормонально-метаболических процессов у маловесных к сроку гестации детей. Педиатрия. Восточная Европа. 2015; 1: 130-41. [Sukalo A.V., Prilutskaya V.A., Solntseva A.V., Uvarova E.V. Modern views on the role of adipocytokines in programming hormonal and metabolic processes in small for gestational age children. Pediatrics. Eastern Europe. 2015; 1: 130-41 (in Russian)].
- Mendez-Ramirez F., Barbosa-Sabanero G., Romero-Gutierrez G., Malacara J.M. Ghrelin in small-for-gestational-age babies: a cross-sectional study. Clin. Endocrinol. (Oxf). 2009; 70(1): 41-6. https://dx.doi.org/10.1111/j.1365-2265.2008.03278.x
- Han L., Li B., Xu X., Liu S., Li Z., Li M., Wang D. Umbilical cord blood adiponectin, leptin, insulin, and ghrelin in premature infants and their association with birth outcomes. Front. Endocrinol. (Lausanne). 2021; 12: 738964. https://dx.doi.org/10.3389/fendo.2021.738964
- Министерство здравоохранения Российской Федерации. Клинические рекомендации. Недостаточный рост плода, требующий предоставления медицинской помощи матери (задержка роста плода). 2025. [Ministry of Health of the Russian Federation. Clinical guidelines (treatment protocol). Insufficient fetal growth requiring medical care for the mother (fetal growth retardation). 2025 (in Russian)].
- Papageorghiou A.T., Ohuma E.O., Altman D.G., Todros T., Cheikh Ismail L., Lambert A. et al. International standards for fetal growth based on serial ultrasound measurements: the fetal growth longitudinal study of the INTERGROWTH-21st project. Lancet. 2014; 384(9946): 869-79. https://dx.doi.org/10.1016/S0140-6736(14)61490-2
- Рюмина И.И., Байбарина Е.Н., Нароган М.В., Маркелова М.М., Орловская И.В., Зубков В.В., Дегтярев Д.Н. Использование международных стандартов роста для оценки физического развития новорожденных и недоношенных детей. Неонатология: новости, мнения, обучение. 2023; 11(2): 48-52. [Ryumina I.I., Baibarina E.N., Narogan M.V., Markelova M.M., Orlovskaya I.V., Zubkov V.V., Degtyarev D.N. Using international growth standards to assess the physical development of newborns and premature infants. Neonatology: News, Opinions, Training. 2023; 11(2): 48-52 (in Russian)]. https://dx.doi.org/10.33029/2308-2402-2023-11-2-48-52
- Кан Н.Е., Солдатова Е.Е., Тютюнник В.Л., Борисова А.Г., Тезиков Ю.В., Липатов И.С., Садекова А.А., Алексеев А.А., Красный А.М. Факторы энергетического метаболизма при задержке роста плода. Акушерство и гинекология. 2024; 5: 44-52. [Kan N.E., Soldatova E.E., Tyutyunnik V.L., Borisova A.G., Tezikov Y.V., Lipatov I.S., Sadekova A.A., Alekseev A.A., Krasnyi A.M. Factors of energy metabolism in fetal growth restriction. Obstetrics and Gynecology. 2024; (5): 44-52 (in Russian)]. https://dx.doi.org/10.18565/aig.2024.9
- Кан Н.Е., Солдатова Е.Е., Тютюнник В.Л., Волочаева М.В., Садекова А.А., Красный А.М. Диагностическая значимость определения экспрессии генов энергетического метаболизма при задержке роста плода. Акушерство и гинекология. 2023; 8: 48-55. [Kan N.E., Soldatova E.E., Tyutyunnik V.L., Volochaeva M.V., Sadekova A.A., Krasnyi A.M. Diagnostic significance of determining the expression of energy metabolism genes in fetal growth retardation. Obstetrics and Gynecology. 2023; (8): 48-55 (in Russian)]. https://dx.doi.org/10.18565/aig.2023.93
- Seferovic M.D., Gupta M.B. Increased umbilical Cord PAI-1 levels in placental insufficiency are associated with fetal hypoxia and angiogenesis. Dis. Markers. 2016; 2016: 7124186. https://dx.doi.org/10.1155/2016/7124186
- Yatsenko T., Us I., Korolova D., Zhuk S., Dziuba H., Nalbat A. et al. Placental dysfunction is associated with dysregulated fibrinolytic system activation. Int. J. Mol. Sci. 2025; 26(19): 9339. https://dx.doi.org/10.3390/ijms26199339
- Bucur-Grosu M.L., Avasiloaiei A., Moscalu M., Dimitriu D.C., Păduraru L., Stamatin M. Desacylated ghrelin and leptin in the cord blood of small-for-gestational-age newborns with intrauterine growth restriction. Acta Endocrinol. (Buchar). 2019; 15(3): 305-10. https://dx.doi.org/10.4183/aeb.2019.305
- Chiesa C., Osborn J.F., Haass C., Natale F., Spinelli M., Scapillati E. et al. Ghrelin, leptin, IGF-1, IGFBP-3, and insulin concentrations at birth: is there a relationship with fetal growth and neonatal anthropometry? Clin. Chem. 2008; 54 (3): 550-8. https://dx.doi.org/10.1373/clinchem.2007.095299
- Napolitano T., Silvano S., Vieira A., Balaji S., Garrido-Utrilla A., Friano M.E. et al. Role of ghrelin in pancreatic development and function. Diabetes Obes. Metab. 2018; 20(Suppl 2): 3-10. https://dx.doi.org/10.1111/dom.13385
- Abdel Hakeem A.H., Saed S.Z., El Rehany M.A., Yassin E.E. Serum level of ghrelin in umbilical cord in small and appropriate for gestational age newborn infants and its relationship to anthropometric measures. J. Clin. Neonatol. 2012; 1(3): 135-8. https://dx.doi.org/10.4103/2249-4847.101694
- James R.J., Drewett R.F., Cheetham T.D. Low cord ghrelin levels in term infants are associated with slow weight gain over the first 3 months of life. J. Clin. Endocrinol. Metab. 2004; 89(8): 3847-50. https://dx.doi.org/10.1210/jc.2003-032167
- Martos-Moreno G.A., Barrios V., Barrios V., Saenz de Pipaon M., Pozo J., Dorronsoro I. et al. Influence of prematurity and growth restriction on the adipokine profile, IGF1, and ghrelin levels in cord blood: relationship with glucose metabolism. Eur. J. Endocrinol. 2009; 161(3): 381-9. https://dx.doi.org/10.1530/EJE-09-0193
- Леонова И. А., Иванов Д. О. Фетальное программирование и ожирение у детей. Детская медицина Северо-Запада. 2015; 6(3): 28-41. [Leonova I.A., Ivanov D.O. Fetal programming and obesity in children. Children's Medicine of the North-West. 2015; 6(3): 28-41 (in Russian)].
- Gonzales-Dominguez M.I., Lazo-de-la-Vega-Monroy M.L., Zaina S., Sabanero M., Daza-Benítez L., Malacara J.M. et al. Ghrelin and GHSR1 receptor in placentas of SGA, LGA and AGA newborns. Endocrine Abstracts. 2015; 37: 5-10. https://dx.doi.org/10.1530/endoabs.37.GP.05.10
- Vickers M.H., Sloboda D.M. Leptin as mediator of the effects of developmental programming. Best Pract. Res. Clin. Endocrinol. Metab. 2012; 26(5): 677-87. https://dx.doi.org/10.1016/j.beem.2012.03.005
- Stefaniak M., Dmoch-Gajzlerska E. Maternal serum and cord blood leptin concentrations at delivery in normal pregnancies and in pregnancies complicated by intrauterine growth restriction. Obes. Facts. 2022; 15(1): 62-9. https://dx.doi.org/10.1159/000519609
- Dodington D.W., Desai H.R., Woo M. JAK/STAT - emerging players in metabolism. Trends Endocrinol. Metab. 2018; 29(1): 55-65. https://dx.doi.org/10.1016/j.tem.2017.11.001
- Barker D.J. The origins of the developmental origins theory. J. Intern. Med. 2007; 261(5): 412-7. https://dx.doi.org/10.1111/j.1365-2796.2007.01809.x
- Pekal Y., Özhan B., Enli Y., Özdemir Ö.M.A., Ergin H. Cord blood levels of spexin, leptin, and visfatin in term infants born small, appropriate, and large for gestational age and their association with newborn anthropometric measurements. J. Clin. Res. Pediatr. Endocrinol. 2022; 14(4): 444-52. https://dx.doi.org/10.4274/jcrpe.galenos.2022.2022-4-24
Received 09.12.2025
Accepted 09.04.2026
About the Authors
Natalia E. Kan, Professor, Dr. Med. Sci., Honored Scientist of the Russian Federation, Deputy Director General for Research – Director of the Institute of Obstetrics, Academician V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology, Ministry of Health of Russia, 117997, Russia, Moscow,Ac. Oparina str., 4, kan-med@mail.ru, Researcher ID: B-2370-2015, SPIN: 5378-8437, Authors ID: 624900, Scopus Author ID: 57008835600,
https://orcid.org/0000-0001-5087-5946
Victor L. Tyutyunnik, Professor, Dr. Med. Sci., Honored Doctor of the Russian Federation, Leading Researcher at the Center of Scientific and Clinical Researches, Academician V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology, Ministry of Health of Russia, 117997, Russia, Moscow, Ac. Oparina str., 4, tioutiounnik@mail.ru, Researcher ID: B-2364-2015, SPIN: 1963-1359, Authors ID: 213217, Scopus Author ID: 56190621500, https://orcid.org/0000-0002-5830-5099
Ekaterina E. Soldatova, PhD, Researcher at the Institute of Obstetrics, Academician V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology, Ministry of Health of Russia, 117997, Russia, Moscow, Ac. Oparina str., 4, +7(906)110-51-13, katerina.soldatova95@bk.ru, https://orcid.org/0000-0001-6463-3403
Kristina O. Ryzhova, PhD student, Academician V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology, Ministry of Health of Russia, 117997, Russia, Moscow, Ac. Oparina str., 4, cr.yanina@gmail.com, https://orcid.org/0009-0007-8318-435X
Asiyat R. Gaidarova, PhD, Obstetrician-gynecologist, Researcher at the 1st Obstetric Physiological Department of the Institute of Obstetrics, Academician V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology, Ministry of Health of Russia, 117997, Russia, Moscow, Ac. Oparina str., 4, 95asiya95@mail.ru, https://orcid.org/0000-0003-1415-3318
Aleksey M. Krasnyi, PhD, Head of the Cytology Laboratory, Academician V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology,
Ministry of Health of Russia, 117997, Russia, Moscow, Ac. Oparina str., 4, alexred@list.ru, https://orcid.org/0000-0001-7883-2702
Corresponding author: Ekaterina E. Soldatova, katerina.soldatova95@bk.ru
Similar Articles

