

Vaginal microbiota profile in human papillomavirus-associated cervical disease and its diagnostic value for cervical intraepithelial neoplasia II+
Kepsha M.A., Mezhevitinova E.A., Timofeeva A.V., Fedorov I.S., Donnikov A.E.
Objective. To characterize the vaginal microbiota in patients with different clinical and morphological forms of human papillomavirus (HPV)-associated cervical disease and assess its relationship with the presence of high-grade cervical intraepithelial lesions and cervical cancer (CIN II+).
Materials and methods. This analytical study included 190 women who were examined at the V. I. Kulakov National Medical Research Center for Obstetrics, Gynecology, and Perinatology, Ministry of Health of the Russian Federation. The following groups were defined: NILM/HPV+ (n=50), LSIL (n=52), HSIL (n=52), and cervical cancer (n=36). Quantitative and qualitative assessments of the vaginal microbiota were performed using real-time PCR (Femoflor-16 assay). HPV genotyping and viral load quantification were performed.
Results. Increasing severity of HPV-associated cervical disease was accompanied by a marked reduction in the relative abundance of Lactobacillus spp. and an increase in the proportion of anaerobic microbial communities. The most pronounced dysbiotic changes were observed in patients with cervical cancer (p<0.001). Stratification into CIN I- (NILM/HPV+, LSIL) and CIN II+ (HSIL, cervical cancer) groups revealed statistically significant differences in HPV genotype distribution and the qualitative and quantitative composition of the microbial consortia. Six logistic regression models were developed for the first time, each incorporating three or four independent variables and demonstrating high sensitivity (80–86%) and specificity (75–80%) in identifying CIN II+. These models included various combinations of quantitative characteristics of HPV types 16 and 33 and selected opportunistic pathogens. All models incorporated a novel index proposed by the authors: the ratio of the total quantities of "possibly carcinogenic" HPV types to "carcinogenic" HPV types, which increased model sensitivity by 4–18%.
Conclusion. Distinct vaginal microbiota profiles are associated with severe HPV-related cervical disease. A comprehensive assessment of the vaginal microbiome in conjunction with HPV status may serve as an additional tool for differentiating clinically significant cervical lesions.
Authors' contributions. Kepsha M.A. – data collection, literature search and analysis, drafting of the manuscript; Mezhevitinova E.A. – data collection, editing of the manuscript, approval of the final version of the manuscript; Timofeeva A.V. – drafting of the manuscript, approval of the final version of the manuscript; Fedorov I.S. – statistical analysis; Donnikov A.E. – obtaining experimental data.
Conflicts of interest. The authors have no conflicts of interest to declare.
Funding. There was no funding for this study.
Ethical Approval. The study was reviewed and approved by the Research Ethics Committee of the V.I. Kulakov NMRC for OG&P, Ministry of Health of Russia.
Patient Consent for Publication. All patients provided informed consent for the publication of their data.
Authors' Data Sharing Statement. The data supporting the findings of this study are available upon request from the corresponding author after approval from the principal investigator.
For citation: Kepsha M.A., Mezhevitinova E.A., Timofeeva A.V., Fedorov I.S., Donnikov A.E. Vaginal microbiota profile in human papillomavirus-associated cervical disease and its diagnostic value for cervical intraepithelial neoplasia II+.
Akusherstvo i Ginekologiya/Obstetrics and Gynecology. 2026; (5): 108-120 (in Russian)
https://dx.doi.org/10.18565/aig.2026.71
Keywords
human papillomavirus (HPV)
vaginal microbiota
cervical intraepithelial neoplasia
CIN I-
CIN II+
Cervical cancer is a major global health challenge. According to the International Agency for Research on Cancer, 662,301 new cases and 348,874 deaths were recorded in 2022 [1]. Carcinogenic HPV genotypes are recognized as necessary etiological factors in the development of cervical cancer [2]. A causal association with invasive cervical cancer has been established for 17 HPV genotypes, with HPV 16 (61.7%) and 18 (15.3%) accounting for the largest proportion of cases. The combined attributable fraction of these genotypes was lowest in Africa and highest in Central, Western, and Southern Asia, whereas the contribution of HPV 35 was significantly higher in Africa than in other regions [3].
The development of precancerous cervical lesions is associated with persistent HPV infections. However, the mechanisms underlying viral clearance in some women and its long-term persistence in others remain insufficiently understood.
According to international studies, low-grade squamous intraepithelial lesions (LSIL), corresponding to cervical intraepithelial neoplasia grade I (CIN I), undergo spontaneous regression in 60% of cases, persist in 30%, and progress to high-grade lesions (HSIL; CIN II–III and carcinoma in situ) in 10%, with invasive cervical cancer developing in only 1% of cases [4].
Accumulating evidence indicates a role for the microbiota in the development and progression of malignancies at various anatomical sites, leading to the emergence of the «oncobiome» concept in the contemporary scientific literature [5, 6]. In HPV-mediated carcinogenesis, disturbances in the lower genital tract microbiota have been linked to unfavorable infection outcomes. Dysbiotic changes, characterized by reduced abundance of Lactobacillus spp. and increased prevalence of opportunistic pathogens (OPs), including Gardnerella vaginalis, are associated with a higher risk of HPV persistence and progression of cervical squamous intraepithelial lesions [7].
A 2019 meta-analysis of 15 observational studies involving more than 100,000 women examined the association between vaginal microbiota composition, HPV status, and severity of cervical lesions. Vaginal dysbiosis was associated with an increased risk of HPV infection (overall OR 1.33; 95% CI 1.18–1.5; I²=0%), with a stronger association observed in younger women (OR 1.43; 95% CI 1.1–1.85; I²=0%). Dysbiosis was also linked to an increased likelihood of persistent HPV infection (overall OR 1.14; 95% CI 1.01–1.28; I²=44.2%), including high-risk types (OR 1.18; 95% CI 1.01–1.38; I²=0%), and an elevated risk of HSIL and invasive cervical cancer (OR 1.18; 95% CI 1.01–1.38; I²=0%) [8].
The literature also suggests that vaginal microbiota influences local immune mechanisms, potentially promoting either viral clearance or persistence [9, 10]. The dominance of specific Lactobacillus species, particularly Lactobacillus crispatus, is considered a protective factor, as it is associated with a stable vaginal microbiome characterized by low microbial diversity and more favorable clinical outcomes. A prospective study demonstrated that the predominance of Lactobacillus crispatus in patients with CIN II was associated with a higher likelihood of lesion regression at 12 months, whereas reduced abundance of lactobacilli and increased microbial diversity were linked to a lack of regression, including progression to CIN III [11].
In this context, a detailed characterization of the composition and quantitative features of the vaginal microbiota associated with either HPV persistence or protective effects is of considerable scientific and clinical interest.
This study aimed to characterize the vaginal microbiota in patients with different clinical and morphological forms of HPV-associated cervical disease and assess its association with high-grade cervical intraepithelial neoplasia and cervical cancer (CIN II+).
Materials and methods
A total of 190 women were examined in an analytical study with prospective group allocation at the outpatient research department of V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology. Eligible participants were women who underwent evaluation for cervical pathology. The inclusion criteria were as follows: availability of cytological and/or histological results, presence of high-risk HPV infection, absence of antibacterial, antiviral, and/or immunomodulatory therapy within 30 days prior to sampling, and written informed consent.
At baseline, the patients underwent cervical cytology (liquid-based or conventional), HPV testing of cervical epithelial scrapings, quantitative analysis of the vaginal microbiota using the Femoflor-16 assay (DNA-Technology LLC), and extended colposcopy. The HPV Kvant-21 assay was used for the detection, genotyping, and quantification of HPV DNA, including types HPV 6, 11, 16, 18, 51, 56, 68, 31, 33, 35, 39, 45, 52, 58, 59, 26, 53, 66, 82, 76, and 44(55). According to the International Agency for Research on Cancer classification, HPV genotypes 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, and 59 are classified as carcinogenic (Group 1); HPV 68 as probably carcinogenic (Group 2A); HPV 53, 66, and 73 as possibly carcinogenic (Group 2B); and HPV 6, 11, and 44(55) as unclassifiable regarding carcinogenicity in humans (Group 3) [12].
Quantitative and qualitative microbiota analysis included assessment of the following microorganisms: Lactobacillus spp., Enterobacteriaceae spp., Streptococcus spp., Staphylococcus spp., Gardnerella vaginalis, Prevotella, Porphyromonas spp., Eubacterium spp., Sneathia spp., Leptotrichia spp., Fusobacterium spp., Megasphaera spp., Veillonella spp., Dialister spp., Lachnobacterium spp., Corynebacterium spp., Mobiluncus spp., Peptostreptococcus spp., Fannyhessea vaginae (formerly Atopobium vaginae), Ureaplasma urealyticum, Ureaplasma parvum, Mycoplasma hominis, Candida spp., and Mycoplasma genitalium. Absolute normocenosis was defined as Lactobacillus spp. comprising >80% of the total bacterial mass (TBM). A relative abundance of 20–80% was considered moderate dysbiosis, and <20% was defined as severe dysbiosis.
Based on the initial findings, a cervical biopsy with subsequent histological examination was performed to verify HPV-associated cervical pathology. Four clinical and morphological groups were established: NILM/HPV+ (n=50), LSIL (n=52), HSIL (n=52), and cervical cancer (n=36).
Statistical analysis
Statistical analyses were performed using scripts written in R [13] and RStudio [14]. The normality of the data distribution was assessed using the Shapiro–Wilk test. For non-normally distributed data, comparisons between independent groups were conducted using the Mann–Whitney U-test. Data are reported as median (Me) with first and third quartiles (Q1; Q3). The significance threshold was set at p=0.05, and values of p<0.001 were reported accordingly. Categorical variables are presented as absolute counts and percentages, and comparisons were performed using Fisher’s exact test.
Logistic regression was used to develop predictive models for the occurrence of CIN II+. The model performance was evaluated using receiver operating characteristic (ROC) analysis, which illustrates the relationship between the true-positive and false-positive rates. Optimal cutoff points were determined to achieve the best discrimination between the groups. Logistic regression models (linear regression with logit transformation) were developed in RStudio using stepwise inclusion and exclusion of microbiological predictors based on their contribution to the model. The diagnostic performance was assessed using the area under the ROC curve (AUC), statistical significance, sensitivity, and specificity.
Results
To enable clinically meaningful stratification and improve interpretability, patients were categorized into two principal groups: CIN I- (NILM/HPV+, LSIL) and CIN II+ (HSIL, cervical cancer), consistent with the current concepts of the biological and prognostic continuum of HPV-associated cervical lesions.
The CIN I- group was characterized predominantly by a transient course of papillomavirus infection, a high likelihood of spontaneous regression, and, in most cases, no indication of active therapeutic intervention. In contrast, the CIN II+ group was defined by the persistence of oncogenic HPV types and an increased risk of progression, necessitating active treatment strategies and heightened oncologic vigilance.
In the first stage, a statistical analysis of clinical variables (patient age, body mass index, age at sexual debut, number of sexual partners at the time of the study, parity, history of hormonal contraception use, HPV vaccination status, history of sexually transmitted infections, smoking status, history of recurrent vulvovaginitis, and chronic diseases) was performed in the defined group. Statistically significant intergroup differences were identified for the following variables: patient age (p<0.001), body mass index (p=0.016), age at sexual debut (p<0.001), number of sexual partners at the time of the study (p<0.001), parity (p<0.001), and history of hormonal contraception use (p=0.023).
Subsequently, a comparative analysis of the proportion of Lactobacillus spp. within the total bacterial mass (TBM) was conducted in patients with CIN I- and CIN II+ to assess the differences in the vaginal microbiota according to lesion severity. In women with CIN I-, normocenosis predominated, with a high relative abundance of Lactobacillus spp. in most cases. In contrast, the CIN II+ group demonstrated a significant reduction in the proportion of Lactobacillus spp. (p=0.004), accompanied by marked interindividual variability, indicating a predominance of dysbiosis.
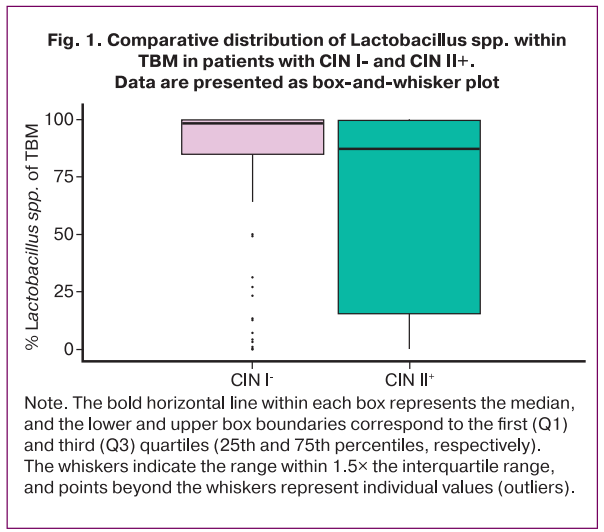
The bold horizontal line within each box represents the median, and the lower and upper box boundaries correspond to the first (Q1) and third (Q3) quartiles (25th and 75th percentiles, respectively). The whiskers indicate the range within 1.5× the interquartile range, and points beyond the whiskers represent individual values (outliers).
These findings indicate that more severe cervical lesions are associated with disruption of the vaginal microbiota, characterized by a reduced relative abundance of Lactobacillus spp. and the development of dysbiotic profiles.
Next, an intergroup comparison of the microbiological characteristics was performed. Statistically significant differences in the prevalence of specific HPV genotypes were identified: HPV types 53 and 66 predominated in the CIN I- group, whereas HPV types 16 and 45 were most frequent in the CIN II+ group (Table 1). These findings reflect differences in oncogenic potential and their contributions to clinically significant lesions.
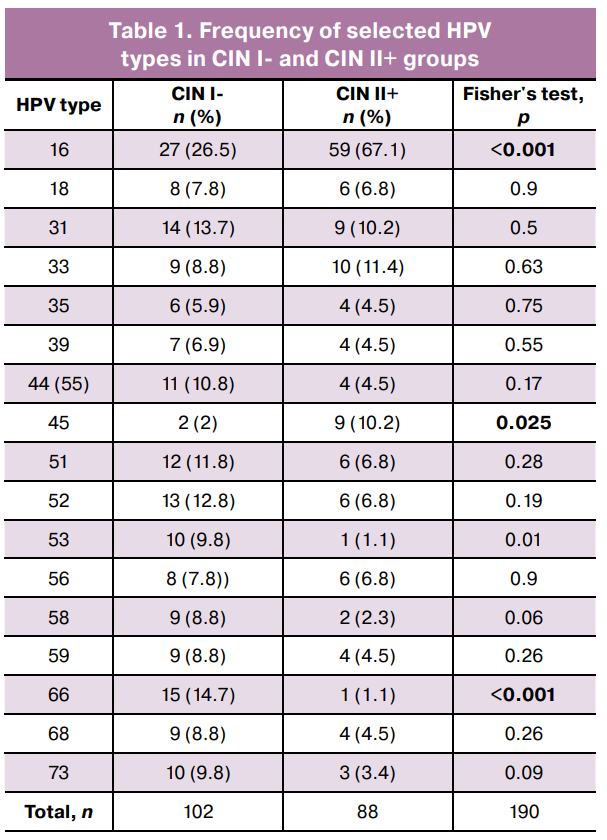
Additionally, the common logarithm of the ratio of opportunistic pathogen (OPs) copy numbers to Lactobacillus spp. was calculated for both groups (Table 2). This metric enabled the assessment of the contribution of individual microorganisms to dysbiosis and its association with dyplasia development.
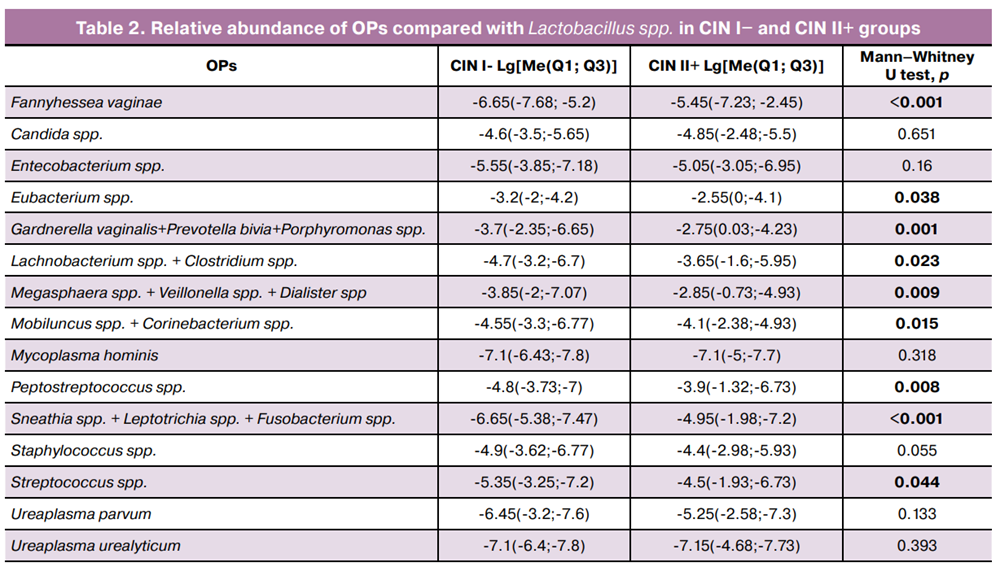
The OP-to-Lactobacillus spp. ratio revealed a statistically significant increase in the relative abundance of several anaerobic bacteria in the CIN II+ group compared that to in the CIN I- group. The most pronounced differences were observed for Fannyhessea vaginae, Eubacterium spp., Gardnerella vaginalis + Prevotella bivia + Porphyromonas spp., and Lachnobacterium spp. + Clostridium spp., Megasphaera spp. + Veillonella spp. + Dialister spp., Mobiluncus spp. + Corinebacterium spp., Peptostreptococcus spp., Sneathia spp. + Leptotrichia spp. + Fusobacterium spp., and Streptococcus spp.. These findings indicate that increased copy numbers of specific OPs in the context of reduced Lactobacillus spp. are associated with more severe cervical lesions.
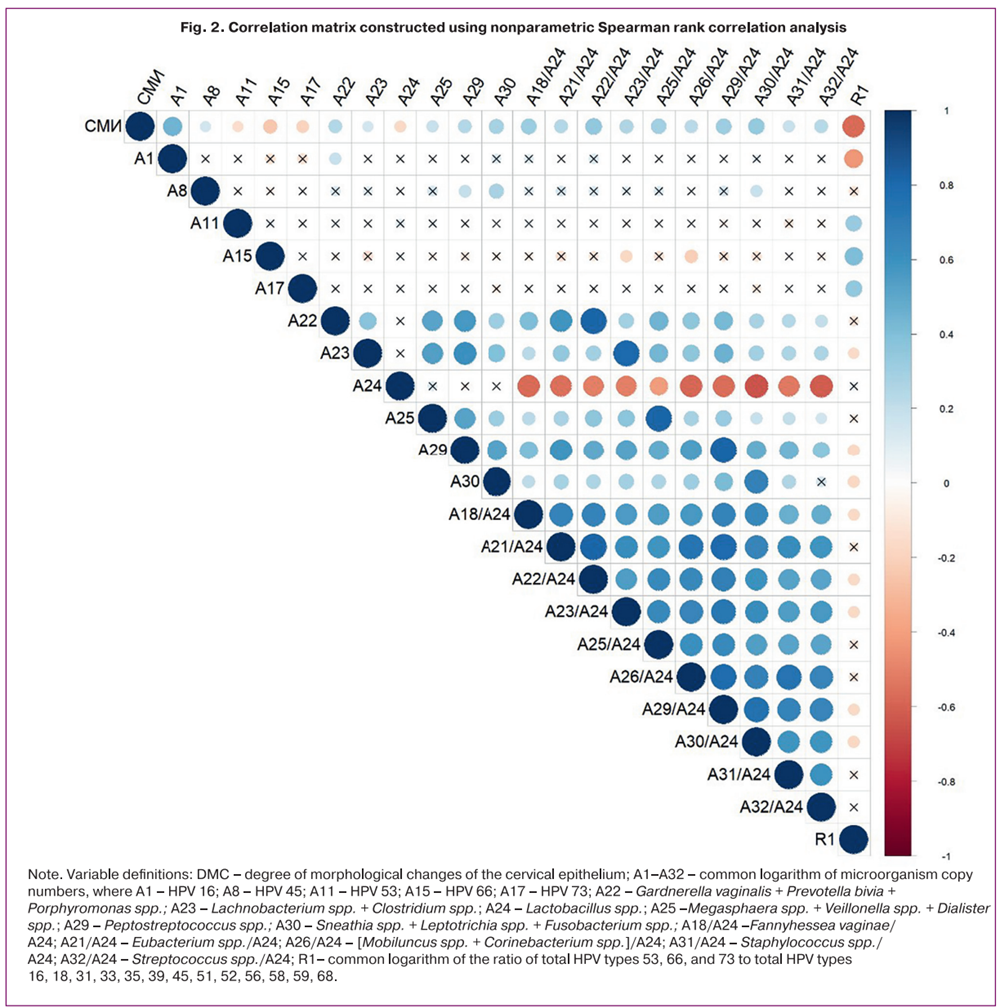
Spearman’s correlation analysis was performed to evaluate the relationship between lesion severity, HPV genotype, and vaginal microbiota parameters (Fig. 2). Statistically significant positive correlations (r≥0.3; p<0.05) were identified between the degree of morphological changes (DMC), as determined by cytological and/or histological examination, and the copy numbers of HPV type 16, Fannyhessea vaginae/Lactobacillus spp., [Sneathia spp. + Leptotrichia spp. + Fusobacterium spp.]/Lactobacillus spp., [Gardnerella vaginalis + Prevotella bivia + Porphyromonas spp.]/Lactobacillus spp., and Peptostreptococcus spp./Lactobacillus spp. The DMC grading was defined as follows: 1 – NILM/HPV+, 2 – LSIL, 3 – HSIL, 4 – cervical cancer.
Notably, significant inverse correlations were observed between DMC and the copy numbers of «possibly carcinogenic» HPV types: HPV 53 (r=-0.15, p=0.03), HPV 66 (r=-0.25, p<0.001), HPV 73 (r=-0.2, p=0.007); the ratio R1 (the common logarithm of the ratio of the total copy number of HPV types 53, 66, and 73 to that of «probably carcinogenic» and «carcinogenic» HPV types 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68) (r=-0.6, p<0.001); and the level of Lactobacillus spp. (r=-0.17, p=0.02).
Importantly, the strength of the positive associations between DMC and Gardnerella vaginalis, Prevotella bivia, Porphyromonas spp., and Lachnobacterium spp. + Clostridium spp., Megasphaera spp. + Veillonella spp. + Dialister spp., Peptostreptococcus spp., and Sneathia spp. + Leptotrichia spp. + Fusobacterium spp. increased when these variables were expressed relative to Lactobacillus spp. (r=0.24 vs r=0.34; r=0.15 vs r=0.25; r=0.18 vs r=0.29; r=0.24 vs r=0.30; r=0.28 vs r=0.33, respectively), consistent with the statistically significant intergroup differences shown in Table 2.
Note. Variable definitions: DMC – degree of morphological changes of the cervical epithelium; A1–A32 – common logarithm of microorganism copy numbers, where A1 – HPV 16; A8 – HPV 45; A11 – HPV 53; A15 – HPV 66; A17 – HPV 73; A22 – Gardnerella vaginalis + Prevotella bivia + Porphyromonas spp.; A23 – Lachnobacterium spp. + Clostridium spp.; A24 – Lactobacillus spp.; A25 –Megasphaera spp. + Veillonella spp. + Dialister spp.; A29 – Peptostreptococcus spp.; A30 – Sneathia spp. + Leptotrichia spp. + Fusobacterium spp.; A18/A24 –Fannyhessea vaginae/A24; A21/A24 – Eubacterium spp./A24; A26/A24 – [Mobiluncus spp. + Corinebacterium spp.]/A24; A31/A24 – Staphylococcus spp./ A24; A32/A24 – Streptococcus spp./A24; R1– common logarithm of the ratio of total HPV types 53, 66, and 73 to total HPV types 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68.
Based on the identified correlations, univariate regression analysis was performed to select predictors for evaluating the association between viral and bacterial factors and the severity of HPV-associated cervical pathology (Table 3). For univariate regression, a statistical significance threshold of p<0.25 was applied for inclusion in the multivariable models [15, 16]. The dependent variable was binary (0 – CIN I-; 1 – CIN II+), and the independent variables comprised the microbiological parameters of the lower genital tract. A total of 22 variables met the inclusion criteria (Table 3).
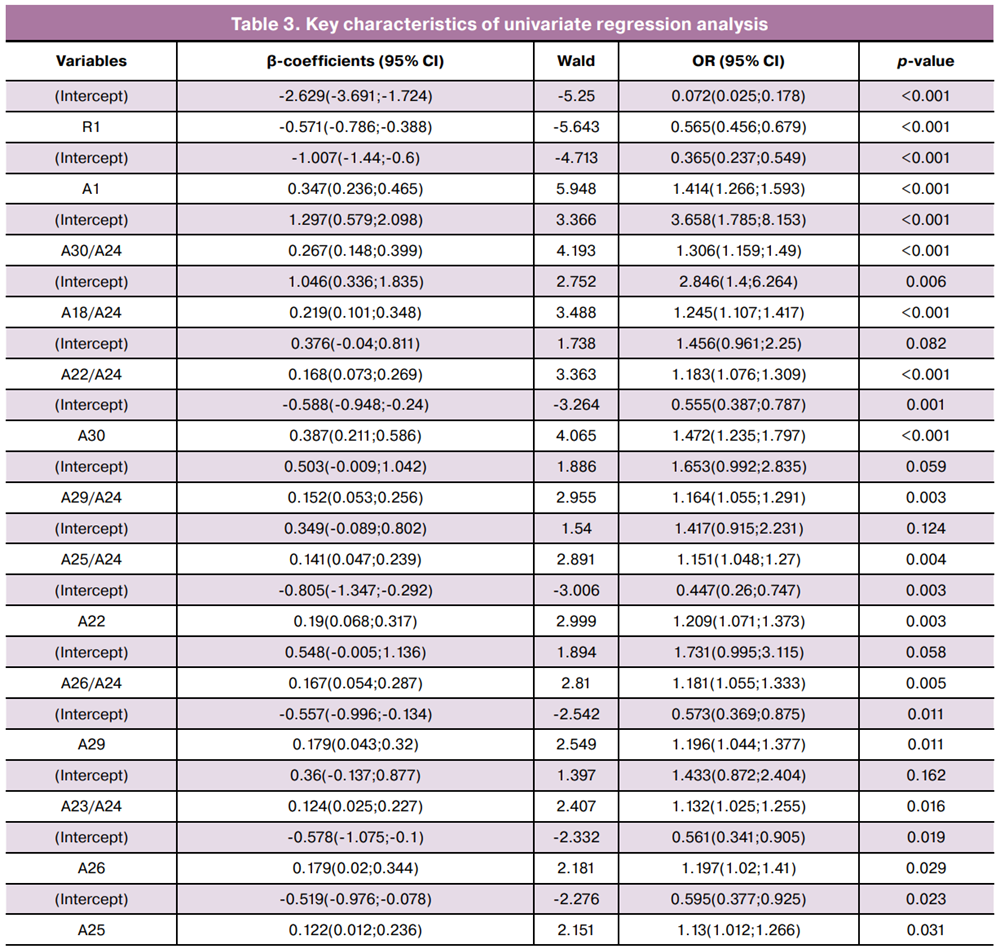
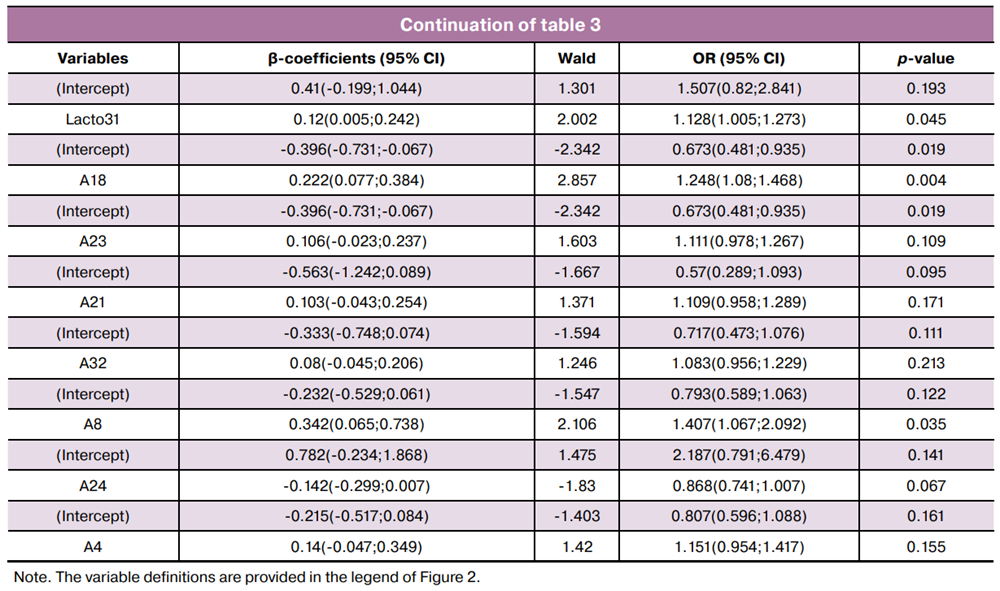
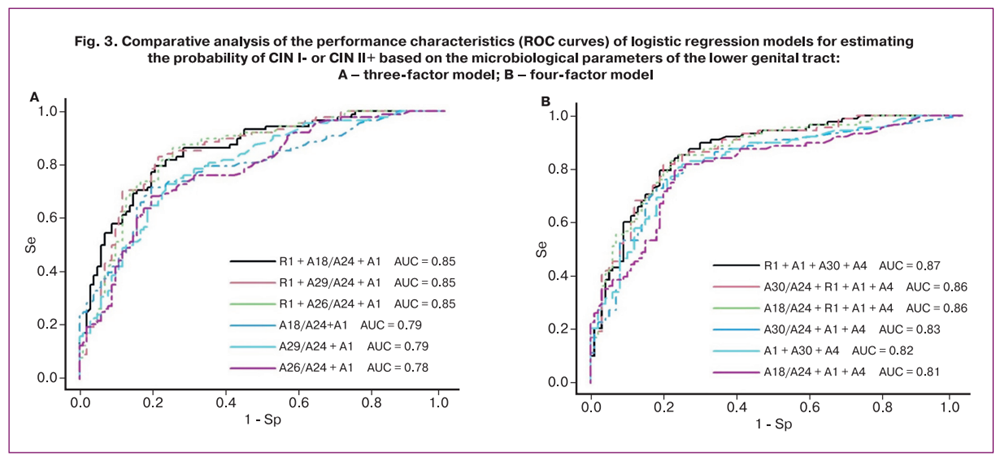
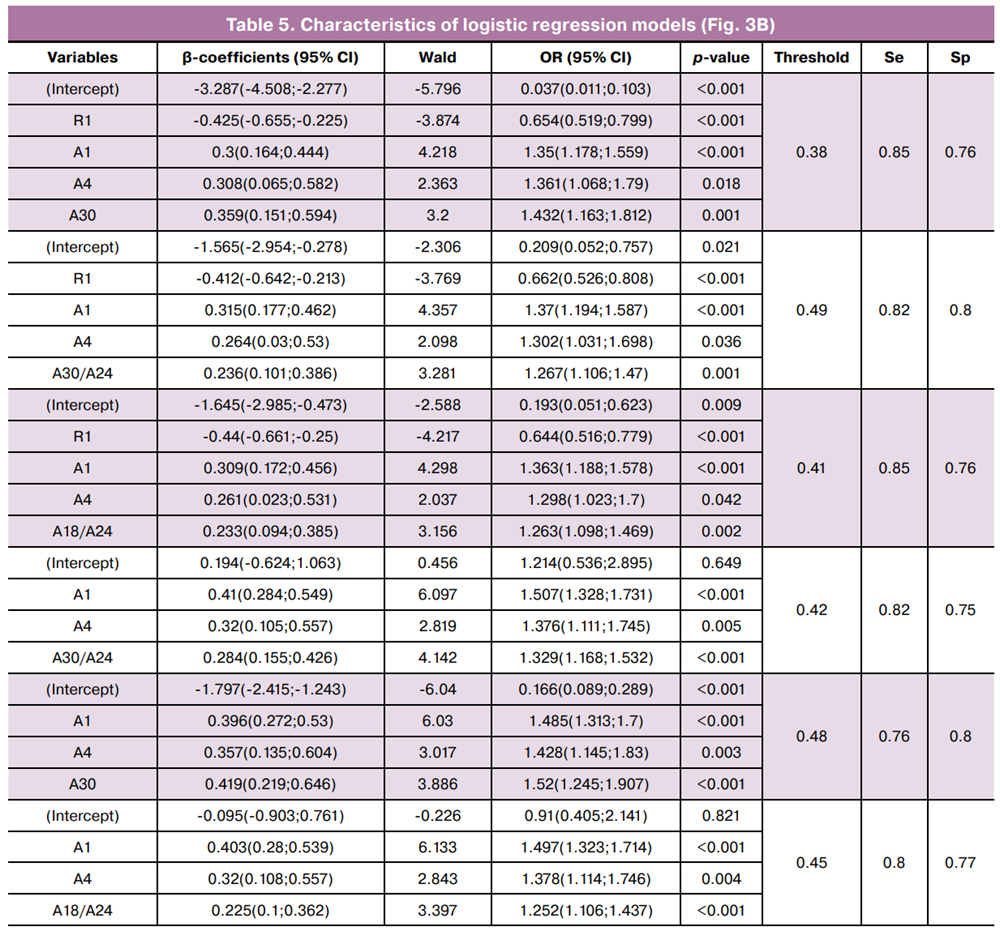
The required sample size for multivariable modeling was calculated using the formula N=50+8n, where n is the number of predictor variables [17]. Thus, for a sample of 190 women, the maximum allowable number of predictors was 17. Using RStudio, stepwise inclusion/exclusion of the 22 selected variables was performed to construct ROC curves based on three (Fig. 3A) and four-variable (Fig. 3B) models.
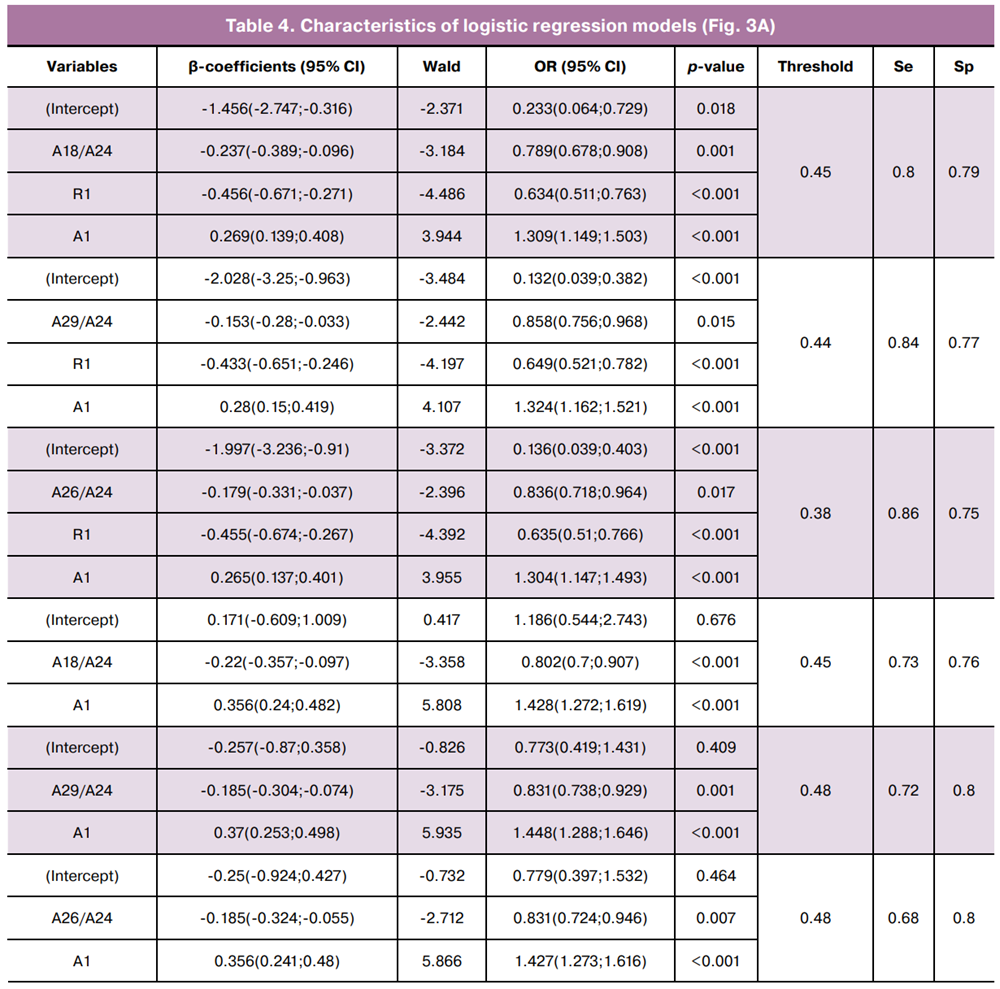
The equations describing the first three logistic regression models selected for practical application are as follows:
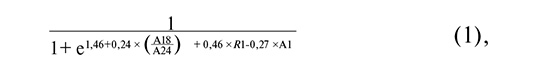
where A18/A24 is the common logarithm of the ratio of Fannyhessea vaginae to Lactobacillus spp. copy numbers; R1 is the common logarithm of the ratio of total HPV types 53, 66, and 73 to total HPV types 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68; A1 is the common logarithm of HPV 16 copy number.

where A29/A24 is the common logarithm of the ratio of Peptostreptococcus spp. to Lactobacillus spp. copy numbers; R1 is as defined above; A1 is the common logarithm of HPV 16 copy number.
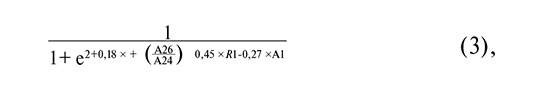
where A26/A24 is the common logarithm of the ratio of [Mobiluncus spp. + Corinebacterium spp.] to Lactobacillus spp. copy numbers; R1 is as defined above, and A1 is the common logarithm of the HPV 16 copy number.
As shown in Figure 3A and Table 4, the inclusion of R1 as an independent variable increased the area under the curve (AUC) from 0.78–0.79 to 0.85 and improved sensitivity from 68–73% to 80–86%.
The equations describing the first three four-variable logistic regression models selected for practical application are as follows:
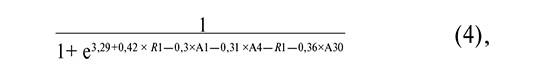
where R1 is as defined above, A1 is the common logarithm of the HPV 16 copy number, A4 is the common logarithm of the HPV 33 copy number, and A30 is the common logarithm of the total Sneathia spp. copy number. + Leptotrichia spp. + Fusobacterium spp.
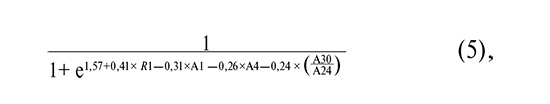
where R1 is as defined above, A1 is the common logarithm of the HPV 16 copy number, A4 is the common logarithm of the HPV 33 copy number, and A30/A24 is the common logarithm of the ratio of [Sneathia spp. + Leptotrichia spp. + Fusobacterium spp.] to Lactobacillus spp.
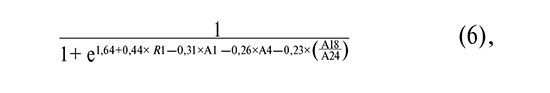
where R1 is as defined above; A1 is the common logarithm of the HPV 16 copy number; A4 is the common logarithm of the HPV 33 copy number; and A18/A24 is the common logarithm of the ratio of Fannyhessea vaginae to Lactobacillus spp.
As shown in Figure 3B and Table 5, the inclusion of R1 increased the AUC from 0.81–0.83 to 0.86–0.87 and improved the sensitivity from 76–82% to 82–85%.
Overall, the models presented in Figures 3A and 3B demonstrated high specificity (75–80%) and sensitivity (80–86%), corresponding to a false positive rate (overdiagnosis of CIN II+) of 20–25% and a false negative rate of 14–20%.
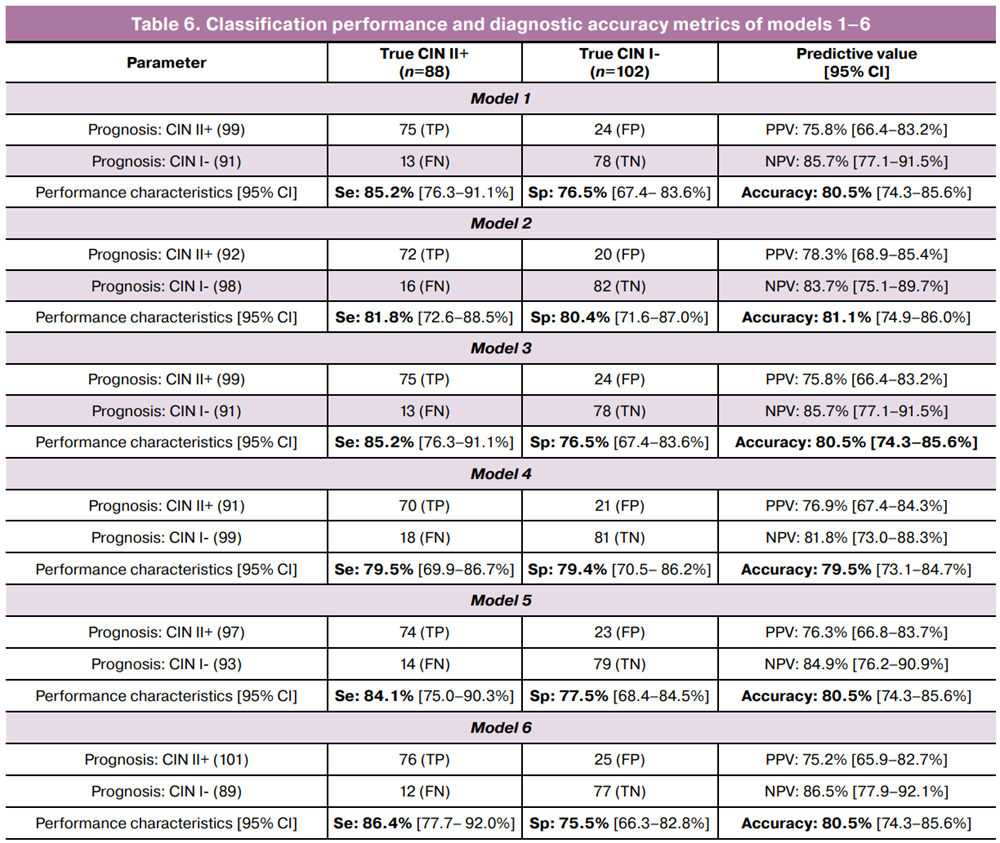
To evaluate the performance of the models selected for further validation and potential clinical application, contingency tables were constructed (Table 6), including sensitivity, specificity, negative predictive value (NPV), and positive predictive value (PPV).
Discussion
The causal relationships between alterations in vaginal microbiota and HPV persistence have not yet been conclusively established. It remains debated whether disruptions in the vaginal microbial community precede viral acquisition and persistence, or whether they arise secondarily as a consequence of virus-induced changes in epithelial integrity and local immunity. Nevertheless, pronounced dysbiotic states – including bacterial vaginosis (BV) – are recognized as factors associated with an increased likelihood of HPV infection, prolonged viral persistence, and progression of cervical lesions.
The present study demonstrates a sequential restructuring of the vaginal microbiome as HPV-associated cervical pathology advances—from morphologically intact epithelium to invasive cervical cancer. This restructuring is characterized by a progressive decline in the proportion of Lactobacillus spp. and a concurrent increase in the relative abundance of opportunistic pathogenic taxa and consortia. This microbial profile is potentially associated with elevated vaginal pH, reduced colonization resistance, and the establishment of a pro-inflammatory microenvironment, collectively creating conditions unfavorable for HPV clearance and potentially facilitating cervical carcinogenesis [18].
The statistically significant associations identified by correlation analysis between clinico-morphological groups, HPV genotypes, and vaginal microbiome parameters reflect the complex viral-microbial profile reorganization accompanying the progression of HPV-associated cervical lesions. These findings are consistent with evidence from international studies.
Lin W. et al. demonstrated a decline in the relative abundance of Lactobacillus spp. and a concomitant increase in anaerobic microorganisms – including Fannyhessea vaginae, Megasphaera spp., Sneathia spp., and Fusobacterium spp. –with increasing severity of cervical lesions [19]. Comparable conclusions were reported in the systematic review and meta-analysis by Norenhag J. et al., which described BV-like anaerobic profiles in CIN II–III, involving Gardnerella vaginalis, Fannyhessea vaginae, Prevotella bivia, Sneathia spp., Megasphaera spp., and Fusobacterium spp. [20]. The association between vaginal dysbiosis and HPV infection has been corroborated by large-scale meta-analyses. Based on 13 studies, Liang Y. et al. [21] demonstrated that BV is associated with an increased risk of HPV infection (OR=2.57; 95% CI 1.78–3.71; p<0.05) and the development of cervical intraepithelial neoplasia. Yang J. et al. [22] confirmed that BV increases the risk of HPV infection 2.45-fold (95% CI 1.83–3.27; p<0.00001). A meta-analysis encompassing 19 studies identified a statistically significant association between BV and CIN (OR=1.51; 95% CI 1.24–1.83) [23].
Of particular note is the negative correlation identified between HPV genotypes 53, 66, and 73 and the severity of morphological changes. Although the International Agency for Research on Cancer classifies HPV genotypes 53, 66, and 73 as «possibly carcinogenic» (Group 2B) [12], their contribution to the development of CIN II+ appears to be less pronounced compared with high-risk genotypes. Several studies have shown that these genotypes are more frequently detected in low-grade cervical lesions and transient HPV infection, consistent with the findings of the present study and potentially reflecting genotype-specific characteristics of viral biological behavior [24, 25].
For the first time, six logistic regression models were constructed to differentiate between the CIN I- and CIN II+ clinico-morphological groups. These models incorporated various combinations of quantitative parameters for HPV type 16, HPV type 33, specific opportunistic microorganisms, and the ratio of the total copy numbers of «possibly carcinogenic» to «carcinogenic» HPV types. The models demonstrated high specificity (75–80%) and sensitivity (80–86%), underscoring their potential clinical utility for the differentiation of clinically significant cervical lesions. It should be noted that the proposed models require external validation in independent cohorts, as well as assessment of the reproducibility of their diagnostic performance across different clinical settings.
HPV is an epitheliotropic virus whose life cycle is intimately linked to epithelial differentiation: primary infection occurs in proliferating basal layer cells, while productive replication and virion assembly take place in differentiating suprabasal layers [26]. Given the epitheliotropic nature of HPV, its dependence on epithelial differentiation throughout its life cycle, and the anatomical and histological heterogeneity of the cervix (ectocervix, endocervix, and transformation zone), HPV-associated disease may exhibit a multifocal pattern. Contemporary studies indicate that a single patient may harbor multiple HPV genotypes, with discrete lesion foci associated with specific viral types, reflecting the heterogeneity of the disease process and the differential oncogenic potential of individual HPV genotypes [27, 28].
In this context, it appears warranted to standardize specimen collection procedures for cervical cytology and HPV testing, with mandatory sampling of the transformation zone. Furthermore, incorporating the ratio of «possibly carcinogenic» types (53, 66, and 73) to the total number of «probably carcinogenic» (68) and carcinogenic (16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59) HPV types, as well as the ratio of copy numbers of specific opportunistic microorganisms to Lactobacillus spp., may enhance the diagnostic value of these assessments.
In summary
- Progressive severity of HPV-associated cervical pathology is accompanied by a restructuring of the vaginal microbiome, characterized by a decline in Lactobacillus spp. and an increase in the relative abundance of anaerobic opportunistic microorganisms.
- CIN II+ lesions are distinguished by a more pronounced predominance of opportunistic microorganisms over Lactobacillus spp. compared with the CIN I- group.
- Genotype-specific differences among HPV types were identified. High-risk genotypes – most notably HPV 16 and 45 – were associated with more severe lesions, whereas genotypes 53, 66, and 73 were more frequently detected in cases with less advanced morphological changes.
- Logistic regression models developed on the basis of quantitative HPV and vaginal microbiome parameters demonstrated high sensitivity and specificity for the diagnosis of CIN II+.
- Comprehensive assessment of HPV status and vaginal microbiome parameters may serve as an adjunctive tool for stratifying patients with HPV-associated cervical pathology.
Conclusion
The present study identified a shift in the composition of the vaginal microbiome toward dysbiosis with increasing severity of HPV-associated cervical lesions. The virus-microbiome associations revealed herein enabled the development of logistic regression models that accurately differentiate between CIN I- and CIN II+. These findings support the value of comprehensive, integrated assessment of HPV status and vaginal microbiome parameters in patients with HPV-associated cervical pathology.
References
- International Agency for Research on Cancer. Absolute numbers of cervical cancer incidence and mortality in 2022. Available at: https://www.iarc.who.int/cancer-type/cervical-cancer/
- Jouya S., Shahabinia Z., Mazidimoradi A., Allahqoli L., Salehiniya H., Lee D.Y. Cervical cancer epidemiology: global incidence, mortality, survival, risk factors, and equity in HPV screening and vaccination. J. Clin. Med. 2026; 15(3): 1079. https://dx.doi.org/10.3390/jcm15031079
- Wei F., Georges D., Man I., Baussano I., Clifford G.M. Causal attribution of human papillomavirus genotypes to invasive cervical cancer worldwide: a systematic analysis of the global literature. Lancet. 2024; 404(10451): 435-44. https://dx.doi.org/10.1016/S0140-6736(24)01097-3
- Zhang X., Bai J., Yuan C., Long L., Zheng Z., Wang Q. et al. Bioinformatics analysis and identification of potential genes related to pathogenesis of cervical intraepithelial neoplasia. J. Cancer. 2020; 11(8): 2150-7. https://dx.doi.org/10.7150/jca.38211
- Dhar R., Kumar A., Karmakar S. OncoBiome: the emerging frontier in cancer-microbiome interactions. Asian J. Med. Sci. 2025; 16(7): 1-2. https://dx.doi.org/10.71152/ajms.v16i7.4629
- Zitvogel L., Derosa L., Routy B., Loibl S., Heinzerling L., de Vries I.J.M. et al. Impact of the ONCOBIOME network in cancer microbiome research. Nat. Med. 2025; 31(4): 1085-98. https://dx.doi.org/10.1038/s41591-025-03608-8
- Mitra A., MacIntyre D.A., Marchesi J.R., Lee Y.S., Bennett P.R., Kyrgiou M. The vaginal microbiota, human papillomavirus infection and cervical intraepithelial neoplasia: what do we know and where are we going next? Microbiome. 2016; 4(1): 58. https://dx.doi.org/10.1186/s40168-016-0203-0
- Brusselaers N., Shrestha S., van de Wijgert J., Verstraelen H. Vaginal dysbiosis and the risk of human papillomavirus and cervical cancer: systematic review and meta-analysis. Am. J. Obstet. Gynecol. 2019; 221(1): 9-18. https://dx.doi.org/10.1016/j.ajog.2018.12.011
- Castanheira C.P., Sallas M.L., Nunes R.A.L., Lorenzi N.P.C., Termini L. Microbiome and cervical cancer. Pathobiology. 2020; 88(2): 187-97. https://dx.doi.org/10.1159/000511477
- Zhou Z.W., Long H.Z., Cheng Y., Luo H.Y., Wen D.D., Gao L.C. From microbiome to inflammation: the key drivers of cervical cancer. Front. Microbiol. 2021; 12(FEB): 767931. https://dx.doi.org/10.3389/fmicb.2021.767931
- Mitra A., MacIntyre D.A., Ntritsos G., Smith A., Tsilidis K.K., Marchesi J.R. et al. The vaginal microbiota associates with the regression of untreated cervical intraepithelial neoplasia 2 lesions. Nat. Commun. 2020; 11(1): 1999. https://dx.doi.org/10.1038/s41467-020-15856-y
- International Agency for Research on Cancer. Agents classified by the IARC monographs. Volumes 1-140. Lyon: IARC; 2026. Available at: https://monographs.iarc.who.int/agents-classified-by-the-iarc/
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at: https://www.r-project.org (accessed on Mar 10, 2021).
- RStudio Team. RStudio: Integrated Development for R. RStudio, Inc., Boston, MA; 2016. Available at: http://www.rstudio.com (accessed on Mar 23, 2021).
- Hosmer D.W., Lemeshow S., Sturdivant R.X. Applied Logistic Regression. Third Edition. Wiley; 2013. https://dx.doi.org/10.1002/9781118548387
- Bursac Z., Gauss C.H., Williams D.K., Hosmer D.W. Purposeful selection of variables in logistic regression. Source Code Biol. Med. 2008; 3: 17. https://dx.doi.org/10.1186/1751-0473-3-17
- Green S.B. How many subjects does it take to do a regression analysis. Multivariate Behav. Res. 1991; 26(3): 499-510. https://dx.doi.org/10.1207/s15327906mbr2603_7
- Shen J., Sun H., Chu J., Gong X., Liu X. Cervicovaginal microbiota: a promising direction for prevention and treatment in cervical cancer. Infect. Agent. Cancer. 2024; 19(1): 13. https://dx.doi.org/10.1186/s13027-024-00573-8
- Lin W., Zhang Q., Chen Y., Dong B., Xue H., Lei H. et al. Changes of the vaginal microbiota in HPV infection and cervical intraepithelial neoplasia: a cross-sectional analysis. Sci. Rep. 2022; 12(1): 2812. https://dx.doi.org/10.1038/s41598-022-06731-5
- Norenhag J., Du J., Olovsson M., Verstraelen H., Engstrand L., Brusselaers N. The vaginal microbiota, human papillomavirus and cervical dysplasia: a systematic review and network meta-analysis. BJOG. 2020; 127(2): 171-80. https://dx.doi.org/10.1111/1471-0528.15854
- Liang Y., Chen M., Qin L., Wan B., Wang H. A meta-analysis of the relationship between vaginal microecology, human papillomavirus infection and cervical intraepithelial neoplasia. Infect. Agent Cancer. 2019; 14: 29. https://dx.doi.org/10.1186/s13027-019-0243-8
- Yang J., Long X., Li S., Zhou M., Hu L.N. The correlation between vaginal pathogens and high-risk human papilloma virus infection: a meta-analysis of case-control studies. Front. Oncol. 2024; 14: 1423118. https://dx.doi.org/10.3389/fonc.2024.1423118
- Gillet E., Meys J.F.A., Verstraelen H., Verhelst R., De Sutter P., Temmerman M. et al. Association between bacterial vaginosis and cervical intraepithelial neoplasia: systematic review and meta-analysis. PLoS One. 2012; 7(10): e45201. https://dx.doi.org/10.1371/journal.pone.0045201
- So K.A., Lee I.H., Lee K.H., Hong S.R., Kim Y.J., Seo H.H. et al. Human papillomavirus genotype-specific risk in cervical carcinogenesis. J. Gynecol. Oncol. 2019; 30(4): e52. https://dx.doi.org/10.3802/jgo.2019.30.e52
- Zhang J., Cheng K., Wang Z. Prevalence and distribution of human papillomavirus genotypes in cervical intraepithelial neoplasia in China: a meta-analysis. Arch. Gynecol. Obstet. 2020; 302(6): 1329-37. https://dx.doi.org/10.1007/s00404-020-05787-w
- Graham S. Keratinocyte differentiation-dependent human papillomavirus gene regulation. Viruses. 2017; 9(9): 245. https://dx.doi.org/10.3390/v9090245
- Bruno M.T., Scalia G., Cassaro N., Boemi S. Multiple HPV 16 infection with two strains: a possible marker of neoplastic progression. BMC Cancer. 2020; 20(1): 444. https://dx.doi.org/10.1186/s12885-020-06946-7
- Awadalla M., Al Rawi H.Z., Alahmadi R.M., Khojah O.T., Al-Shouli S.T., Almansour M.I. et al. Transcriptomic profile of the immune genes, oncogenes, and tumor suppressor genes in HPV associated Cervical Intraepithelial Neoplasia 3 (CIN 3) and Cervical Squamous Cell Carcinoma (CSCC): Comparable expressions indicative of invasive potential. Tumour Virus Res. 2025; 20: 200327. https://dx.doi.org/10.1016/j.tvr.2025.200327
Received 27.02.2026
Accepted 10.04.2026
About the Authors
Maria A. Kepsha, Junior Researcher at the Scientific and Outpatient Department, V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology, Ministry of Health of Russia, 4, Ac. Oparin str., Moscow, 117997, Russia, m_kepsha@oparina4.ru, https://orcid.org/0000-0002-4201-1360Elena A. Mezhevitinova, Dr. Med. Sci., Leading Researcher at the Scientific and Outpatient Department, V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology, Ministry of Health of Russia, 4, Ac. Oparin str., Moscow, 117997, Russia, mejevitinova@mail.ru, https://orcid.org/0000-0003-2977-9065
Angelica V. Timofeeva, PhD (Bio), Head of the Laboratory of Applied Transcriptomics of the Department of Systems Biology in Reproduction, Institute of Translational Medicine, V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology, Ministry of Health of Russia, 4, Ac. Oparin str., Moscow, 117997, Russia, avtimofeeva28@gmail.com, https://orcid.org/0000-0003-2324-9653
Ivan S. Fedorov, Researcher at the Laboratory of Transcriptomic, Department of Systems Biology in Reproduction, Institute of Translational Medicine, V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology, Ministry of Health of Russia, 4, Ac. Oparin str., Moscow, 117997, Russia, is_fedorov@oparina4.ru, https://orcid.org/0000-0002-2104-5887
Andrey E. Donnikov, PhD, Head of the Laboratory of Molecular Genetic Methods, V.I. Kulakov National Medical Research Center for Obstetrics, Gynecology and Perinatology, Ministry of Health of Russia, 4, Ac. Oparin str., Moscow, 117997, Russia, donnikov@dna-technology.ru, https://orcid.org/0000-0003-3504-2406
Similar Articles

