
Возрастающий интерес к развитию вспомогательных репродуктивных технологий (ВРТ)
во всем мире обусловлен приоритетностью проблемы бесплодия. Современные подходы к диагностике и лечению бесплодных супружеских пар основаны на достижениях фундаментальной
науки в области изучения молекулярно-генетических процессов, лежащих в основе реализации
процесса репродукции у человека.
Способность эмбриона к успешной имплантации определяется различными факторами, зависит от условий формирования исходного статуса гамет, стабильности организации их генетического материала, строгой временнóй последовательности структурных внутриклеточных процессов [4, 7].
С тех пор как S.M. Wykes и соавт. в 1997 г. первыми опубликовали данные о существовании
нескольких видов мРНК в зрелом сперматозоиде человека, отцовский вклад в оплодотворение
и развитие эмбриона стал важным предметом исследований [5, 11, 19].
В течение последних 15 лет определены различные подгруппы популяций мРНК. Транскриптом
сперматозоида содержит более 400 видов мРНК, роль которых в процессе сперматогенеза и раннего эмбриогенеза остается недостаточно изученной [4, 5]. Имеются данные, что сперматозоид
может передавать эти мРНК ооциту при оплодотворении, что доказано на моделях мышей
и у человека [4, 5, 14]. В литературе также есть указания на то, что спермальные мРНК обладают структурной функцией в связи с тем, что они составляют часть ядерного матрикса, располагаясь по периферии ядра близко к ядерной мембране [7, 14, 15].
В процессе сперматогенеза происходит ремоделирование хроматина. На постмейотической
гаплоидной стадии сперматогенеза происходит замещение гистонов и гистоноподобных белков,
упаковывающих ДНК протаминами через ряд транзиторных белков [1, 2]. Протамины принадлежат к небольшим ядерным протеинам.
В результате участия протаминов в организации ядерного матрикса головки сперматозоида хроматин становится транскрипционно неактивным вследствие высокой конденсации ДНК. В работах последних лет была показана связь между количеством транскриптов протаминов, концентрацией протаминов, подвижностью сперматозоидов и способностью к оплодотворению [5,
10, 11, 17]. Дефицит протаминов может являться результатом дисфункции транскрипции и трансляции в течение сперматогенеза [4].
Немаловажным для исхода программ ВРТ является также пенетрационная способность сперматозоида. Ключевую роль в процессе взаимодействия сперматозоида и ооцита играют белки семей ства
интегральных мембранных протеинов ADAM (A Disintegrin And Metalloproteinase), известных также
как фертилин-α (ADAM-1) и фертилин-β (ADAM-2) [8, 13, 18]. Эти белки локализуются в экваториальной области головки сперматозоида, а растворимые домены фертилинов могут связываться с микроворсинчатым регионом ооцита. Учитывая данные британских исследователей о том, что у человека ген фертилина-α неактивен и является псевдогеном, предметом нашего изучения явилась экспрессия мРНК фертилина-β [9]. Фертилин-β может потенцировать связывание головки сперматозоида и способствовать слиянию гамет [3]. Однако окончательная роль и вклад фертилина-β в формирование зиготы и развитие эмбриона до сих пор остаются неизвестными.
Таким образом, профиль мРНК может иметь значение для развития сперматозоида и раннего
эмбриогенеза. Литературные данные о геномном и эпигеномном наследовании у человека
позволяют сделать заключение о потенциальной важности «отцовского вклада» не только для
оплодотворения ооцита, но и для раннего развития эмбриона, а также возможности использования этих данных для усовершенствования программ ВРТ. Разработка неинвазивных методов для определения качества сперматозоидов в прогнозировании развития эмбриона с хорошим имплантационным потенциалом позволит оптимизировать программы ВРТ.
В связи с этим цель настоящего исследования состояла в определении связи между уровнями экспрессии мРНК протаминов 1 и 2 типов (PRM-1 и PRM-2), фертилина-β (ADAM-2) и исходами лечения бесплодия при использовании ВРТ.
Материал и методы исследования
С целью определения взаимосвязи уровней экспрессии спермальных мРНК генов PRM-1,
PRM-2, ADAM-2 и исхода программ ВРТ было проведено исследование спермы у 79 мужчин,
включенных в исследование и участвующих в программе экстракорпорального оплодотворения
и переноса эмбриона (ЭКО и ПЭ). Критериями включения пар в исследование служили: трубно-перитонеальный фактор бесплодия, возраст пациентки не более 37 лет, число предшествующих циклов ЭКО и ПЭ не более 2, нормальный овариальный резерв. Критериями исключения
явились – наличие уровня фолликулостимулирующего гормона в начале менструального цикла
более 12 мМЕ/мл, эндометриоз II–IV степеней распространения, миома матки больших размеров,
синдром поликистозных яичников, олигоастенотератозооспермия III–IV степеней. Стимуляция
суперовуляции проводилась по стандартному «длинному» протоколу. Перенос 1 или 2 эмбрионов в полость матки проводился на 3-и и/или 5-е сут дробления.
Разделение исследуемых пациентов на группы было произведено с учетом данных европейской группы мониторинга исходов ВРТ (EIM ESHRE, 2007). Были выделены две группы супружеских
пар успешного (I группа) и неуспешного исходов программы ЭКО и ПЭ (II группа). В I группу (контроля) были включены пациенты с благополучным результатом лечения бесплодия, что привело к рождению здорового ребенка (n=13). Исследуемые пациенты с неуспешным завершением ЭКО и ПЭ были распределены в 3 подгруппы. У пациенток подгруппы IIa беременность наступила, но закончилась самопроизвольным выкидышем до 11 нед (n=10). Подгруппу IIb составили пациентки с биохимической беременностью, не подтвержденной далее ультразвуковыми данными (уровень β-субъединицы хорионического гонадотропина составлял не менее 30 мМЕ/мл на 14-й
день после переноса эмбрионов), так называемые «преэмбрионические потери» (n=19). К IIс подгруппе были отнесены пациентки с отрицательным исходом ЭКО и ПЭ, уровень β-субъединицы
хорионического гонадотропина составлял менее 20 мМЕ/мл (n=37). Всего беременность наступила
у 23 пациенток из 79 исследуемых пар. Частота наступления клинической беременности в расчете
на ПЭ составила 29,1%.
Для исследования образцы нативной спермы были отобраны в день трансвагинальной пункции в отдельные пробирки в объеме 200 мкл. Исследование профиля мРНК генов PRM-1, PRM-2 и ADAM-2 проводили методом количественной полимеразной цепной реакции с обратной транскрипцией. Уровень экспрессии оценивали методом ΔΔCq, сравнивая нормированные
значения пороговых циклов исследуемых генов. Нормировку проводили относительно референсных генов HPRT1, TBP и B2М. За 1 было принято значение медианы (Ме) в контрольной группе.
При статистической обработке данных в качестве меры центральной тенденции количественных признаков выбрана медиана, а в качестве интервальной оценки – верхнюю и нижнюю
квартили. Для оценки значимости межгрупповых различий применялся U-критерий Манна-
Уитни.
Результаты исследования
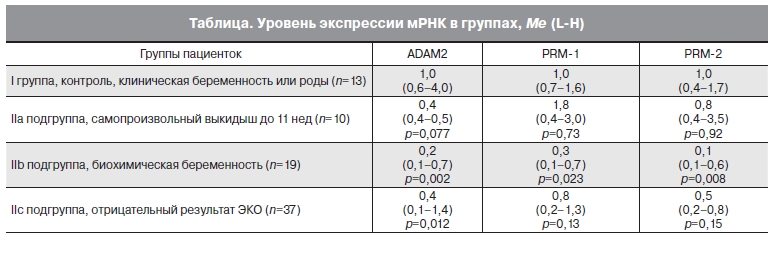
В результате проведенного исследования было показано, что при неуспешном исходе программы ЭКО и ПЭ определялось достоверное снижение уровня экспрессии мРНК фертилина-β (ADAM-2) в нативной сперме. При этом в подгруппе IIb уровень снизился в 6,3 раза (Me=0,16, p=0,002), а в подгруппе IIc – в 2,6 раза (Ме=0,38, p=0,012) относительно контрольной группы (табл. 1). Аналогичная тенденция к снижению уровня экспрессии ADAM-2 прослеживалась и в IIa подгруппе – в 2,2 раза по сравнению с контрольной группой (р=0,077).
Таблица. Уровень экспрессии мРНК в группах, Me (L-H).
При анализе различий уровней экспрессии мРНК фертилина-β между контрольной группой
и II группой в целом, выявлено статистически значимое снижение экспрессии фертилина-β
в 2,6 раза в группе II (p=0,003).
Для мРНК протаминов получены значимые различия между I и IIb группами: уровень экспрессии мРНК гена PRM-1 был в 3,7 раза меньше (p=0,023), а уровень экспрессии мРНК гена
PRM-2 был в 7,1 раза меньше (p=0,008) по сравнению с группой контроля. Аналогичная тенденция
к снижению уровня экспрессии PRM1 и PRM2 прослеживалась и в IIc подгруппе с неуспешным
исходом ЭКО и ПЭ соответственно в 1,2 раза и 1,9 раза по сравнению с контрольной группой
(соответственно р=0,13 и р=0,15). Достоверных различий и тенденции к снижению уровня экспрессии мРНК PRM1 и PRM2 в IIa подгруппе с самопроизвольными выкидышами по сравнению с контрольной группой не выявлено.
Обсуждение
В работе были рассмотрены наиболее важные и хорошо изученные спермальные мРНК
(ADAM-2, PRM-1, PRM-2). ADAM-2, или фертилин-β, играет ключевую роль в слиянии
гамет. Комплексом распознавания фертилина-β служит трипептид FEE (фенилаланин-глутамин-глутамин) в оолемме ооцита [5].
Основная функция протаминов сводится к упаковке и компактизации спермальной ДНК
в сперматозоидах в процессе ремоделирования хроматина. Биологический смысл ремоделирования состоит в том, чтобы отцовский генетический материал был доставлен без риска повреждения молекул ДНК извне, наиболее компактно. В результате конденсации хроматина транскрипционные процессы в сперматозоиде замедляются. Изменение количества протаминов приводит к неадекватной компактизации хроматина, нарушению структурной целостности ДНК. При проведении программ ВРТ сперматозоиды со значительным уровнем фрагментации ДНК сохраняют способность оплодотворять ооциты, однако в дальнейшем эмбриональное развитие может блокироваться на разных этапах.
Степень конденсации хроматина влияет не только на качество спермы, но и на раннее развитие эмбриона. В работах M.J. D’Occhio. и соавт. выявлена значимая корреляция степени конденсации хроматина с выживаемостью эмбриона на ранних стадиях развития [6]. Это может быть связано с тем, что протамины принимают участие во взаимодействии материнского и отцовского геномов и организации упаковки хромосом при первом делении зиготы и последущих первых делениях эмбриона.
Если ранее, до открытия спермальных мРНК, считалось, что качество эмбриона определяется в основном состоянием и степенью «зрелости» ооцита, а роль сперматозоида состоит лишь
в доставке ДНК, то в настоящее время все бóльшее внимание уделяется влиянию мужского фактора
на процессы раннего развития [7, 16, 20]. В настоящее время предполагают, что «отцовский вклад»
в развитие эмбриона состоит не только в доставке и предоставлении генетического материала,
но включает и доставку спермальных мРНК [12]. Активация собственного генома у человеческого
эмбриона происходит, начиная со стадии 8 бластомеров, то есть через 72 ч после оплодотворения. Транскрипты хранятся в цитоплазме задолго до того, как потребуется реализовать процесс трансляции и получить необходимые молекулы белка [12]. Ранее функцию запасания транскриптов для раннего эмбриона отводили исключительно ооциту. Фактически первые 72 ч своего развития эмбрион использует транскрипты, запасенные в гаметах, бóльшей частью – в ооцитах. Демонстрация процесса передачи транскриптов от сперматозоида к ооциту при оплодотворении поддерживает гипотезу об участии спермальных мРНК в развитии зиготы и эмбриона [14].
В результате работы получены данные о негативном влиянии пониженного уровня мРНК протаминов и фертилина-β на исход программы ВРТ, что согласуется с данными M. Depa-Martynow
и соавт. (2007) и B. Kempisty и соавт. (2006) [5, 10]. Однако в нашем исследовании не выявлено столь существенного снижения количества мРНК протаминов и фертилина-β в сперме при неудачах программы ЭКО. По-видимому, это связано с тем, что успешный исход ЭКО и ПЭ
определяется не только изученными факторами, но и другими параметрами, такими как физиологический статус женщины, качество ооцита и переносимых эмбрионов, состояние эндометрия, течение беременности, сопутствующей патологии и др.
Обращает на себя внимание и то, что низкий уровень экспрессии мРНК исследуемых генов
ассоциирован с потерей эмбриона на ранних стадиях развития (биохимическая беременность)
или отсутствием беременности (группы IIb и IIc). Это соответствует современным представлениям
о роли спермальных мРНК в обеспечении основных функций сперматозоида и их участии в раннем эмбриогенезе на преимплантационных и имплантационных стадиях развития. Время полужизни мРНК составляет от 5 до 72 ч, то есть до активации собственного генома эмбриона, достигшего стадии 8 бластомеров. Неблагоприятный прогноз программ ВРТ ассоциирован с уменьшением экспрессии мРНК генов ADAM-2, PRM-1 и PRM-2.
Таким образом, определение экспрессии спермальных мРНК для прогнозирования исхода
программ ВРТ представляется целевым направлением изучения отцовского вклада в развитие
эмбриона. В будущем морфологическая характеристика гамет, скорее всего, будет сопровождаться данными об их молекулярном профиле. Исследование роли спермальных транскриптов в процессе оплодотворения на этапе формирования зиготы и стадии раннего дробления является перспективной областью в углубленной диагностике мужского фактора бесплодия и требует дальнейшего детального анализа.



{kind=link}