
Ежегодно почти у 600 тыс. россиянок выявляются репродуктивные расстройства, восстановление которых, в соответствии с Концепцией демографической политики государства на период до 2025 г., предполагает среди прочих методов использование вспомогательных репродуктивных технологий (ВРТ). Однако применение ВРТ представляет не только новые возможности, но и новые проблемы, что в России связано с отсутствием традиций их практического использования, несмотря на то что многие страны уже накопили почти тридцатилетний опыт (в мире рождены более 4 млн детей с использованием этой технологии). Так, до настоящего времени не сформулированы основные принципы ведения беременности после ВРТ, что связано с отсутствием фундаментальных исследований в этой области.
В настоящее время установлено, что нарушение формирования системы мать-плацента-плод на ранних этапах развития является фактором, во многом определяющим развитие осложнений во время беременности, а в тяжелых случаях предопределяющих материнскую и перинатальную заболеваемость и смертность [1].
С позиций предиктивной медицины пристальное внимание исследователей обращено на генетические особенности системы мать-плацента-плод. Генный полиморфизм факторов, формирующих фетоплацентарную систему, в ряде случаев определяет особенности функционирования этой системы, детерминирует ее реакции на различные воздействия. Наибольший интерес представляют однонуклеотидные генные полиморфизмы (SNP), широко распространенные в популяции и в ряде случаев приводящие к изменению активности гена или модификации белкового продукта. При определенных условиях такие изменения могут приводить к развитию патологических процессов. Выявление соответствующих генных полиморфизмов позволяет выделить группу риска по развитию акушерских осложнений после ВРТ.
Цель исследования: изучить роль генных полиморфизмов, ассоциированных с формированием системы
мать-плацента-плод, в развитии акушерских осложнений у пациенток после ВРТ.
Материал и методы исследования
В исследуемую группу (I группа – индуцированная беременность – ИБ) вошли 167 беременных после применения ВРТ (средний возраст 34,4±4,3 года). Контрольную группу (II группа – самопроизвольная беременность – СБ) составили 145 женщин с самопроизвольной одноплодной беременностью (средний возраст 29,9±4,7 года). Число пациенток, необходимое для исследования, было рассчитано с помощью программы Epi Info версия 6.
В группе ИБ у 69 (41,3%) пациенток беременность была достигнута с помощью традиционного
экстракорпорального оплодотворения, у 57 (34,1%) – с помощью интрацитоплазматической инъекции сперматозоида, а у 41 (24.6%) – после переноса криоконсервированных эмбрионов.
Длительность бесплодного периода в основной группе не превышала 5 лет у 37 (22,1 %), 10 лет – у 81 (48,5%) пациенток, более 10 лет – у 49 (29,4%) пациенток. У 69 (41%) пациенток, беременность наступила с 1-й попытки ВРТ, у 67 (38,9%) пациенток – в анамнезе 1–2 попытки, у 21 (12,5%) пациентки – 3–4 попытки и у 10 (7,6%) пациенток – более 5 попыток. У 109 (65%) пациенток стимуляцию суперовуляции проводили по длинному протоколу и у 46 (35%) – по короткому с антагонистами. Число полученных ооцитов в среднем составило 13,5±5,9, число перенесенных эмбрионов в полость матки – 1,7±0,5.
В большинстве наблюдений бесплодие было вторичным, так, у 41% женщин в анамнезе отмечено до 3 беременностей. Причинами бесплодия являлись: трубно-перитонеальный фактор – 27%, эндометриоз – 21%, синдром поликистозных яичников – 15%, мужской фактор – 23%, бесплодие неясного генеза – 14%.
Из исследования исключали пациенток с многоплодной беременностью, одноплодной беременностью в результате редукции одного эмбриона из двойни, с тяжелой формой экстрагенитальной патологии, с наличием хромосомных аномалий у плода.
В ходе наблюдения за течением беременности пациенткам проводили полный спектр клинико-лабораторного и инструментального обследований.
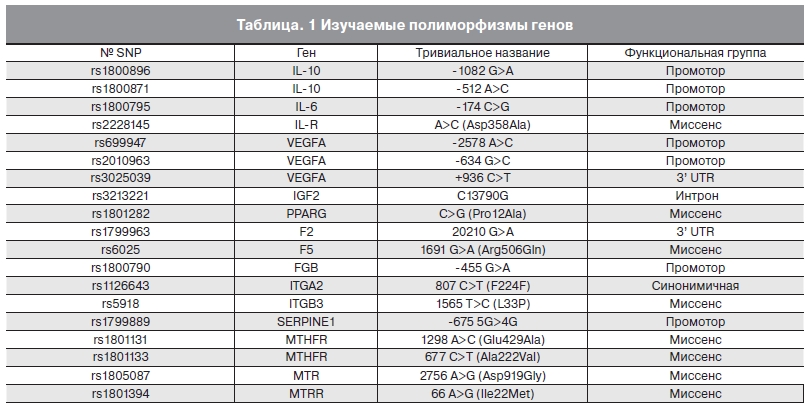
Гены-кандидаты для анализа отбирались на основе их вклада в формирование фето-материнского комплекса. К ним относятся гены, контролирующие ответную реакцию иммунной системы в процессах имплантации и плацентации (IL-10, IL-6, IL-6R), регулирующие формирование сосудистой сети плаценты (VEGFA), метаболические функции плаценты (PPARG, IGF II) (табл. 1).
Таблица.1 Изучаемые полиморфизмы генов.
Выделение ДНК проводили следующим образом: 0,5 мл крови, взятой на EDTA в качестве антикоагулянта, смешивали в 1,5 мл микроцентрифужных пробирках типа Эппендорф с 0,5 мл лизирующего раствора, состоящего из 0,32 М сахарозы, 10 мМ Трис-HCl рН=7,5, 5 мМ MgCl, 1% Тритона Х-100, центрифугировали в течении 1 мин. при 10 000 об/мин, супернатант удаляли, а осадки клеточных ядер два раза отмывали указанным буфером. Последующий протеолиз проводили в 50 мкл буферного раствора, содержащего 50 мМ KCl, 10 мМ Трис-HCl рН=8,3, 2,5 мМ MgCl, 0,45% NP40, 045% Твина 20 и 250 мкг/мл протеиназы К при 37 °С в течение 20 мин. Инактивировали протеиназу К при 98 °С в течение 20 мин. Полученные образцы ДНК до типирования хранили при -20 °С. Концентрация ДНК, определенная на ДНК-минифлуориметре (Ноеfer, США), составляла в среднем 50–100 мкг/мл.
Исследование генного полиморфизма проводили с использованием метода аллель-специфической полимеразной цепной реакции (ПЦР) с последующим гидролизом ампликонов соответствующей рестриктирующей эндонуклеазой. Геномную ДНК из образцов крови выделяли по стандартной методике с использованием протеиназы К, фенол-хлороформной экстракции и осаждения этанолом. Постановку ПЦР проводили в реакционной смеси, содержащей: 5 мкл 10хПЦР-буфера (670 мМ трис-HCl, pH=8,8, 67 mM Mg сульфата аммония, 100 мМ меркаптоэтанола, 10 мкл DMSO, по 12 мМ каждого дНТФ, 10 нг геномной ДНК, 2 единицы Taq-полимеразы и 50 рмоль соответствующей пары праймеров). Идентификацию аллельных вариантов проводили по наличию сайта узнавания для соответствующей рестриктирующей эндонуклеазы с помощью электрофореза в агарозном геле.
Статистический анализ проводили с помощью пакета программ «Statistica for Windows 5.0». Распределение генотипов по исследованным полиморфным локусам проверяли на соответствие равновесию Харди-Вайнберга с помощью точного теста Фишера. Для сравнения частот аллелей между различными группами использовали критерий χ² Пирсона. Различия считали статистически значимыми при р<0,05. Для анализа ассоциации аллелей и генотипов изучаемых генов с риском развития акушерских осложнений рассчитывали отношение шансов (ОШ) с 95% доверительными интервалами (ДИ).
Результаты исследования
Cтруктура акушерских осложнений в исследуемой и основной группах представлена следующим образом. В I триместре беременности кровотечения имели место у 21,6% беременных после ВРТ и 14,3% – при CБ (p=0,033); ранние репродуктивные потери (самопроизвольные выкидыши) – у 14,4% и 8,9% соответственно (p=0,041). Во II и III триместрах отмечены: синдром задержки развития плода – у 14,4% женщин в группе ИБ и 10,1% – в контрольной группе (p=0,025); поздние выкидыши – у 7,5 и 4,2% соответственно (p=0.043); преэклампсия – у 11,7 и 6,5% соответственно (p=0,012); преждевременные роды – у 16,6 и 10,1% соответственно (p=0,037).
Таблица 2. Распределение аллелей и гаплотипов изучаемых генов в исследуемой и контрольной группах.
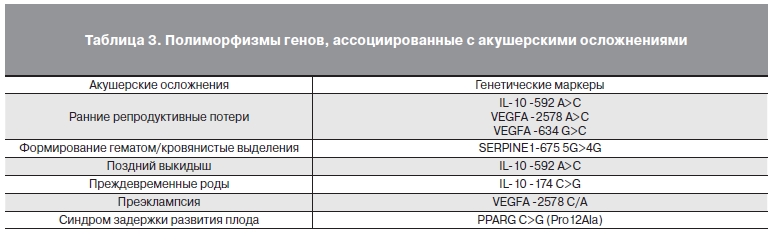
Таким образом, отмечено статистически значимое увеличение частоты акушерских осложнений на протяжении всей беременности в группе женщин, забеременевших после применения ВРТ. В то же время не выявлено статистически значимых различий в распределении аллелей/гаплотипов всех изучаемых генов, ответственных за формирование системы мать-плацента-плод между группой ИБ и СБ (табл. 2) Однако при оценке ассоциации аллелей/генотипов с акушерскими осложнениями установлены статистически значимые различия (табл. 3 и 4).
Таблица 3. Полиморфизмы генов, ассоциированные с акушерскими осложнениями.
Таблица 4. Статистически значимые ассоциации аллелей и генотипов с акушерскими осложнениями в исследуемых группах.
Обсуждение
При носительстве аллеля А полиморфного участка С-592A гена IL-10 в гомозиготном состоянии у пациенток обеих групп увеличивался риск ранних репродуктивных потерь (3,2 и 3,0 соответственно) и поздних выкидышей (1,4 и 1,8 соответственно). Предположительно аллель А усиливает продукцию IL-10 мононуклеарами, что характерно для женщин с привычным невынашиванием беременности. Кроме того, избыточная продукция IL-10 приводит к дисбалансу в иммунной системе, что в свою очередь нарушает экспрессию молекул 1-го и 2-го классов главного комплекса гистосовместимости, при этом в избыточном количестве синтезируются большие гранулоциты и моноциты, что приводит к потере беременности. [11].
Установлено, что IL-6 является важным медиатором в ответе организма на инфекционный агент.
Представленная работа демонстрирует, что гомозиготный генотип GG (замена цитозина на гуанин)
приводит к снижению промоторной активности и в результате к снижению синтеза IL-6, что ассоциировано с риском преждевременных родов как при ИБ, так и при СБ (2,5 и 2,1 соответственно) Полученные результаты согласуются с данными литературы [10].
IL-6 осуществляет свои функции опосредованно через соответствующие рецепторы (IL-6R). Согласно
данным литературы, уровень растворимого IL-6R в амниотической жидкости при преждевременных
родах повышен [8]. D.R. Velez и соавт. [21] обнаружили, что концентрация растворимого рецептора IL-6 в амниотической жидкости строго ассоциирована с гаплотипом IL-6R беременных и плодов, что свидетельствовало о возможности использования системы IL-6/IL-6R в качестве предиктора преждевременных родов. В нашем исследовании не выявлено ассоциации аллелей/генотипов IL-6R A>C (Asp358Ala) с акушерскими осложнениями ни при ИБ, ни при СБ. Система мать-плацента-плод также контролируется генами сосудисто-эндотелиального фактора роста А (VEGFA). VEGFA экспрессируется на материнской и плодовой поверхностях плаценты с ранних сроков беременности, что указывает на его исключительную роль в ангиогенезе трофобласта, дифференциации цитотрофобласта и дальнейшем формировании плаценты. Изменение активации генов VEGFA приводит к нарушениям формирования системы мать-плацента-плод, что в дальнейшем приводит к невынашиванию беременности [7]. Показано, что минорный аллель А промоторной области VEGFA -2578A ассоциирован со снижением экспрессии VEGFA; в отличие от этого полиморфизм -634 G>C увеличивает продукцию VEGFA. В нарушении плацентации играет роль как усиление, так и снижение экспрессии VEGFA. Полученные результаты подтверждаются работами различных авторов. Так, Р. Carmeliet и соавт. установили, что снижение экспрессии VEGFA приводит к гибели эмбрионов [4], в то время как Schneider и соавт. провели исследование, показавшее, что избыточная экспрессия VEGFA также приводит
к невынашиванию беременности [18]. Результаты данного исследования показали, что аллель С полиморфизма гена -2578 A>C ассоциирован с невынашиванием беременности, что согласуется с исследованием D. Papazoglou и соавт. [16]. Кроме того, мы оценивали ассоциацию аллелей/генотипов гена 634 G>C и установили, что носительство С аллеля ассоциировано с ранними репродуктивными потерями в обеих группах (ОШ=1,1 и 1,4, p<0,05), что в целом не противоречит проведенным ранее работам [6]. Кроме того, экспрессию VEGFA может изменять полиморфизм гена +936 C>T. У пациенток, носительниц Т аллеля, отмечается снижение уровня VEGFA, что предрасполагает к развитию преэклампсии [3, 19].
При оценке ассоциации аллелей/генотипов +936 C>T с акушерскими осложнениями не выявлено статистически значимых различий ни при ИБ, ни при СБ, что отличается от результатов, полученных J.Y. Shim и соавт. [19], которые продемонстрировали ассоциацию между полиморфизмом гена +936 C>T и преэклампсией в корейской популяции.
Основной функцией фетоплацентарного комплекса является участие в газовом обмене, метаболизме, поддержании гомеостаза в межворсинчатом пространстве. Уникальностью PPARγ является то, что
он участвует во всех этих процессах [12]. Наиболее изученным полиморфизмом гена PPARG является
полиморфизм C>G (Pro12Ala), при котором происходит замена нуклеотида C на G в 34-м положении экзона B, что приводит к замещению пролина на аланин в аминокислотном положении 12 изоформы PPARγ2 и ассоциировано со снижением активности PPARγ2 [12]. Согласно нашему исследованию, носительство аллеля Ala12 полиморфизма гена PPARG C>G (Pro12Ala) ассоциировано с рождением маловесных детей как при ИБ, так и при СБ (ОШ=2,5 и 2,7; р<0,05), что можно объяснить подавлением активности липаз, участвующих в инсулиновом обмене [12]. Полученные данные подтверждаются работой А. Meirhaeghe и соавт. [12].
Рост и развитие плода находятся по контролем гормонов и факторов роста, важнейшим из которых является семейство инсулиноподобных факторов роста (IGF-I и IGF-II), участвующих в ауто- и паракринной стимуляции пролиферации клеток и их дифференцировки [5, 17]. В опытах на мышах было показано, что их выключение приводит к развитию синдрома Беквита-Видемана [20] или Сильвера-Рассела [9]. В представленном исследовании при оценке ассоциации аллелей/генотипов IGF-II C13790G с акушерскими осложнениями не выявлено статистически значимых различий в обеих группах.
В последние годы в литературе представлено огромное число исследований, посвященных изучению взаимосвязи полиморфизмов генов, регулирующих фолатный обмен, и риска акушерских осложнений. Установлено, что одним из пусковых механизмов является повышение уровня гомоцистеина, что в свою очередь приводит к эндотелиальной клеточной дисфункции и оксидативному стрессу. В нашем исследовании не выявлено ассоциации между акушерскими осложнениями и носительством аллелей полиморфизмов генов-регуляторов ферментов фолатного цикла (677 C>T, 1298 A>C, 2756 A>G, 2756 A>G (Asp919Gly, 66 A>G [Ile22Met]), что можно объяснить высоким потреблением витаминов, содержащих фолаты в исследуемых группах беременных [13, 14].
Среди генетических маркеров предрасположенности к тромбофилии выделяют полиморфизм гена
ингибитора активатора плазминогена 1 типа (PAI-1) [15]. Чрезмерно высокий уровень PAI-1 в эндометрии связывают со снижением глубины инвазии трофобласта и нарушением имплантации [2]. Установлено, что полиморфизм -675 5G/4G гена SERPINE1 связан с делецией в промоторном участке гена и изменяет экспрессию PAI-1. В нашем исследовании мы получили достоверную связь между гетерозиготным гаплотипом -675 5G/4G PAI-1 и кровянистыми выделениями в I триместре беременности в обеих группах. При оценке ассоциаций аллелей/генотипов остальных полиморфизмов, связанных с риском нарушения системы свертывания крови и акушерских осложнений, не выявлено статистически значимых различий ни при ИБ, ни при СБ.
Таким образом, представленная работа подтверждает наличие ассоциаций ряда генных полиморфизмов с акушерскими осложнениями. При этом ни в одном случае не обнаружено статистически значимого влияния способа достижения беременности на силу или направленность ассоциации генетических маркеров с акушерскими осложнениями. Полученные результаты свидетельствуют, что при самопроизвольной или индуцированной беременности действуют одинаковые факторы риска.



{kind=link}
{kind=link}